Studie fra 1926 som forteller at blodets bæringsevne for oksygen er det samme i skilpadder og i mennesker, mens CO2 innholdet er mye høyere hos skilpadder.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2140843/pdf/387.pdf
Studie fra 1926 som forteller at blodets bæringsevne for oksygen er det samme i skilpadder og i mennesker, mens CO2 innholdet er mye høyere hos skilpadder.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2140843/pdf/387.pdf
Svært mye interessant i denne studien om pusten og CO2. Spesielt avsnittene om at kronisk smerte endrer pustemønsteret og senker CO2 nivået i kroppen.
http://www.ncbi.nlm.nih.gov/pubmed/11530714
CHRONIC PAIN
Acute pain results in shortness of breath and an increase in ventila- tion (Nishino, Shimoyama, Ide, & Isono, 1999). A commonly used pain provocation in the laboratory is immersion of a limb into almost freezing water (cold pressor test), which is reliably followed by reduc- tions of PetCO2 among healthy people. (On the other hand, partial or full immersion of the face in cold water causes a modest reduction in ventilation, a component of the diving response). Patients who experi- ence intense chronic pain show these respiratory-related changes over extended periods. For example, migraine headache patients were found to have significantly lowered PetCO2 levels during an attack compared to controls and to migraine-free periods (Hannerz & Jogestrand, 1995), and there were even respiratory abnormalities immediately before an attack (Zhao, Sand, & Sjaastad, 1992). Glynn, Lloyd, & Folkhard (1981) examined arterial pH and PCO2 in 52 chronic pain patients (e.g., back pain, cancer-related pain). PCO2 was mark- edly lowered in these patients, and nerve blockade of pain resulted in a significant rise in PCO2.
Interestingly, blood pH was normal, indicat- ing a long-term blood chemistry compensation for chronic hyperven- tilation. In a sleep study of fibromyalgia patients, a high incidence of respiratory abnormalities such as periodic breathing were found, and arterial PCO2 was lowered in a subgroup of patients (Sergi et al., 1999). Many clinicians, including one of the present authors (Gevirtz), have had the opportunity to measure PetCO2 levels in hun- dreds of chronic muscle pain patients, and the clinical impression is that these levels are almost universally low (c.f., Timmons & Ley, 1994). Of course, pain may also play a role in the increased ventilation found in the FCD patients discussed above, especially during acute episodes of chest pain.
The increased ventilation during acute pain is likely a component of the fight-flight response, preparing the individual for immediate action and sometimes for being attacked or maybe injured. Interest- ingly, recent evidence from animal studies indicates that acute hyperventilation has anesthetic effects via the adrenergic and endogenous opiate system (Ide et al., 1994a, 1994b). Thus, the increased ventila- tion that first served to activate an individual for a fight may have the beneficial side effect of relieving pain if the fight is lost.
So far, no study we know of has examined if the chronic hyperventi- lation exhibited by pain patients is of any benefit to their pain experi- ence (and thus a coping strategy), is only a side effect of the intense pain, or makes their pain worse. One would expect that chronic hyper- ventilation is not healthy in these patients, as it is in other clinical groups, because it interferes with blood homeostatic mechanisms and can lead to a variety of physical symptoms. It has been suggested that by numbing pain, hyperventilation may become a short-term adaptive process with long-term negative consequences (Conway, 1994). Inter- esting in this context is that opioids are frequently prescribed to chronic pain patients to suppress their pain, and they typically also suppress ventilation via central nervous pathways, sometimes to a lethal extent. In summary, there is some initial evidence that hyper- ventilation plays a role in chronic pain, and some mediating mecha- nisms have been identified. However, most of the pain-hypocapnia relationship in chronic pain syndromes is not well understood.
Chronic Pain
Slow abdominal breathing is often taught as a relaxation technique in preparation for acute pain, such as surgery or childbirth, and it also helps patients counteract their tendency to hyperventilate during such events. As described above, the chronic hyperventilation that can accompany long-lasting pain may be especially problematic because it may have long-term negative organismic effects. It is therefore logi- cal that breathing training could be a valuable asset in the overall treat- ment of chronic pain disorders. However, no data are currently avail- able on the role of breathing training as a systematic intervention in these disorders. It is one author’s (Gevirtz) clinical experience that breathing training is in fact a powerful tool in a comprehensive pain management protocol. This is also a common assumption of most bodywork therapies of pain (c.f., Clifton-Smith, 1998). Here again, the capnometry readings are used to illustrate the physiological basis of the symptomatology.
Muscular pain can result from chronically tense muscles. Hubbard, Gevirtz, and their colleagues recently showed that a sympathetically mediated pathway to muscle spindles (trigger points), rather than pathways to muscle fibers, plays an important role in the maintenance of chronic muscular pain (Gerwin, Shannon, Hong, Hubbard, & Gevirtz, 1997; Hubbard & Berkoff, 1993; McNulty, Gevirtz, Hub- bard, & Berkoff, 1994). Psychological stress increased the activity of these spindles, which suggests that stress reduction could alleviate chronic muscle pain. Thus, relaxation induced by slow diaphragmatic breathing may have a beneficial effect on the activation of these spin- dles and reduce general muscle tension.
Full Studie om melatonin sin effetk på mitokondrier.
http://onlinelibrary.wiley.com/doi/10.1111/j.1600-079X.2004.00181.x/full
Abstract: Melatonin, or N-acetyl-5-methoxytryptamine, is a compound derived from tryptophan that is found in all organisms from unicells to vertebrates. This indoleamine may act as a protective agent in disease conditions such as Parkinson’s, Alzheimer’s, aging, sepsis and other disorders including ischemia/reperfusion. In addition, melatonin has been proposed as a drug for the treatment of cancer. These disorders have in common a dysfunction of the apoptotic program. Thus, while defects which reduce apoptotic processes can exaggerate cancer, neurodegenerative disorders and ischemic conditions are made worse by enhanced apoptosis. The mechanism by which melatonin controls cell death is not entirely known. Recently, mitochondria, which are implicated in the intrinsic pathway of apoptosis, have been identified as a target for melatonin actions. It is known that melatonin scavenges oxygen and nitrogen-based reactants generated in mitochondria. This limits the loss of the intramitochondrial glutathione and lowers mitochondrial protein damage, improving electron transport chain (ETC) activity and reducing mtDNA damage. Melatonin also increases the activity of the complex I and complex IV of the ETC, thereby improving mitochondrial respiration and increasing ATP synthesis under normal and stressful conditions. These effects reflect the ability of melatonin to reduce the harmful reduction in the mitochondrial membrane potential that may trigger mitochondrial transition pore (MTP) opening and the apoptotic cascade. In addition, a reported direct action of melatonin in the control of currents through the MTP opens a new perspective in the understanding of the regulation of apoptotic cell death by the indoleamine.
Nevner at et fotbad med CO2 vann gir mer antioksidanter i blodet, mindre fire radikaler og bedre sirkulasjon for de med sirkulasjonsproblemer i føttene.
http://www.ncbi.nlm.nih.gov/pubmed/21248668
The aim of this paper was to investigate the release of oxygen free radicals in patients with peripheral occlusive arterial disease and the effects of immersion of the legs and feet in carbon dioxide (CO(2))-enriched water.
Twenty-five patients with peripheral occlusive arterial disease (Fontaine stage II) and 15 healthy controls were treated by immersing the lower legs in either CO(2)-enriched or normal spa water. Blood samples were collected in heparinized tubes and total antioxidant status (TAS) and reactive oxygen metabolites (ROMs) were measured after five treatments a week for two weeks.
d-ROM plasma levels decreased in patients with peripheral occlusive disease after immersion in CO(2)-enriched water (P<0.001), and in healthy controls (P<0.01), in line with a significant increase in TAS (P<0.001).
CO(2)-enriched water immersion had a positive effect, reducing free radical plasma levels and raising the levels of antioxidants, suggesting an improvement in the microcirculation.
Mye interessant om melatonin her. Bl.a. at det er neuroprotective og hjelper i forsvaret mot oksidativt stress.
http://www.ncbi.nlm.nih.gov/pubmed/11270481
Aerobic cells use oxygen for the production of 90-95% of the total amount of ATP that they use. This amounts to about 40 kg ATP/day in an adult human. The synthesis of ATP via the mitochondrial respiratory chain is the result of electron transport across the electron transport chain coupled to oxidative phosphorylation. Although ideally all the oxygen should be reduced to water by a four-electron reduction reaction driven by the cytochrome oxidase, under normal conditions a small percentage of oxygen may be reduced by one, two, or three electrons only, yielding superoxide anion, hydrogen peroxide, and the hydroxyl radical, respectively. The main radical produced by mitochondria is superoxide anion and the intramitochondrial antioxidant systems should scavenge this radical to avoid oxidative damage, which leads to impaired ATP production. During aging and some neurodegenerative diseases, oxidatively damaged mitochondria are unable to maintain the energy demands of the cell leading to an increased production of free radicals. Both processes, i.e., defective ATP production and increased oxygen radicals, may induce mitochondrial-dependent apoptotic cell death. Melatonin has been reported to exert neuroprotective effects in several experimental and clinical situations involving neurotoxicity and/or excitotoxicity. Additionally, in a series of pathologies in which high production of free radicals is the primary cause of the disease, melatonin is also protective. A common feature in these diseases is the existence of mitochondrial damage due to oxidative stress. The discoveries of new actions of melatonin in mitochondria support a novel mechanism, which explains some of the protective effects of the indoleamine on cell survival.
EPOC er Excess Post-exercise Oxygen Consuption, som innebærer at kroppen bruker mer oksygen etter trening for å fylle på lagrene den har tømt under treningsøkten.
Dette er også en viktig årsak til at pusteteknikker etter treningsøkten har så gode resultater. Ikke minst fordi økt CO2 gjør at O2 lettere hopper av blodcellene og kan benyttes i cellene som trenger det.
EPOC er aktuelt om treningsintensiteten er over 70% VO2max og varer lenger enn 50 minutter, eller er superintens på over 105% VO2max i mer enn 6 minutter.

Wiki siden her: http://en.wikipedia.org/wiki/Excess_post-exercise_oxygen_consumption
In recovery, oxygen (EPOC) is used in the processes that restore the body to a resting state and adapt it to the exercise just performed. These include: hormone balancing, replenishment of fuel stores, cellular repair, innervation and anabolism. Post-exercise oxygen consumption replenishes the phosphagen system. New ATP is synthesized and some of this ATP donates phosphate groups to creatine until ATP and creatine levels are back to resting state levels again. Post-exercise oxygen is also used to oxidize lactic acid. Lactic acid is produced during exercise and then travels via the blood stream to the kidneys, cardiac muscle, and liver. An increased amount of oxygen is necessary to convert the lactic acid back to pyruvic acid at these locations. Another use of EPOC is to fuel the body’s increased metabolism from the increase in body temperature which occurs during exercise.[2]
En studie her: http://www.ncbi.nlm.nih.gov/pubmed/17101527
Recovery from a bout of exercise is associated with an elevation in metabolism referred to as the excess post-exercise oxygen consumption (EPOC).
Evidence has accumulated to suggest an exponential relationship between exercise intensity and the magnitude of the EPOC for specific exercise durations. Furthermore, work at exercise intensities >or=50-60% VO2max stimulate a linear increase in EPOC as exercise duration increases.
it is now apparent that a prolonged EPOC (3-24 h) may result from an appropriate exercise stimulus (submaximal: >or=50 min at >or=70% VO2max; supramaximal: >or=6 min at >or=105% VO2max).
However, even those studies incorporating exercise stimuli resulting in prolonged EPOC durations have identified that the EPOC comprises only 6-15% of the net total oxygen cost of the exercise.
Og en studie til her: http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3527216/
Trained individuals have better thermoregulatory capacities than untrained individuals because physical training enhances the sweating mechanism at a given level of the central sweating drive [11]. Therefore, elevated body temperature in untrained individuals could last longer than in trained individuals [12]. Moreover, subjects with lower VO2max might produce more lactate than those with higher VO2max especially during strenuous exercise. An enhanced lactate metabolism requires oxygen consumption for recovery. Thus, fitness level may contribute to the magnitude of EPOC.
In conclusion, we revealed that cardiorespiratory fitness level correlates inversely with the magnitude of EPOC, especially when performing an aerobic-type interval exercise.
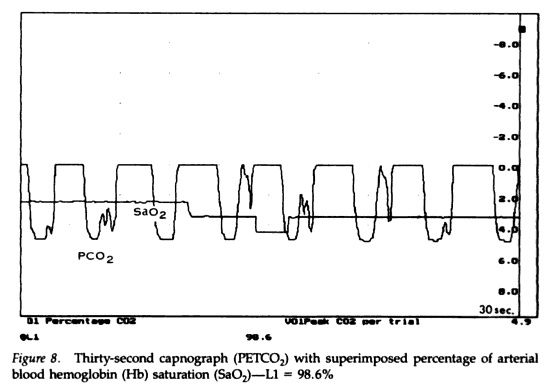
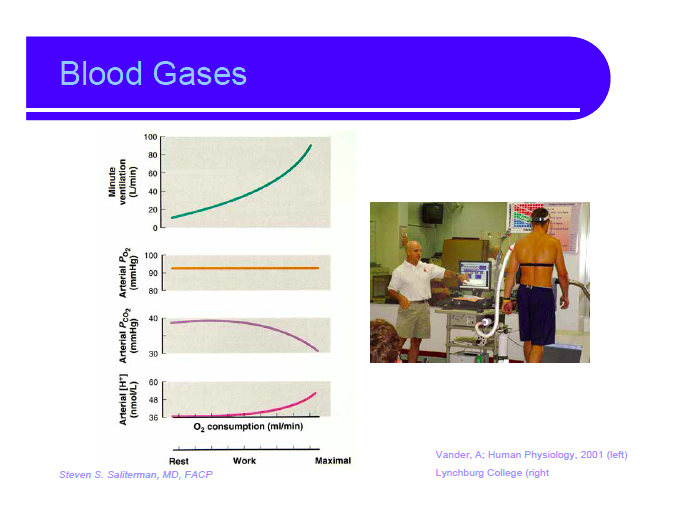
Nevner forholdet mellom CO2 og O2 under hard trening. Har flere gode illustrasjoner om at oksygenforbruket fungerer i hard trening, hvordan blodsirkulasjonen distribueres og hvordan CO2 synker mens O2 metningen er den samme.
http://www.tc.umn.edu/~drsteve/Lectures/Exercise%20Physiology.pdf

One thinks that one is tracing the outline of the thing’s nature over and over again, and one is merely tracing round the frame through which we look at it.
Ludvig Wittgenstein
Omfattende gjennomgang av hvordan kroppen reagerer på CO2. Mange ting om CO2 er nevnt for aller første gang i denne studien fra 2003.
http://www.ncbi.nlm.nih.gov/pubmed/12818238
H(+) is maintained constant in the internal environment at a given body temperature independent of external environment according to Bernard’s principle of «milieu interieur». But CO2 relates to ventilation and H(+) to kidney. Hence, the title of the chapter. In order to do this, sensors for H(+) in the internal environment are needed. The sensor-receptor is CO2/H(+) sensing. The sensor-receptor is coupled to integrate and to maintain the body’s chemical environment at equilibrium. This chapter dwells on this theme of constancy of H(+) of the blood and of the other internal environments. [H(+)] is regulated jointly by respiratory and renal systems.
The respiratory response to [H(+)] originates from the activities of two groups of chemoreceptors in two separate body fluid compartments: (A) carotid and aortic bodies which sense arterial P(O2) and H(+); and (B) the medullary H(+) receptors on the ventrolateral medulla of the central nervous system (CNS). The arterial chemoreceptors function to maintain arterial P(O2) and H(+) constant, and medullary H(+) receptors to maintain H(+) of the brain fluid constant. Any acute change of H(+) in these compartments is taken care of almost instantly by pulmonary ventilation, and slowly by the kidney. This general theme is considered in Section 1.
The general principles involving cellular CO2 reactions mediated by carbonic anhydrase (CA), transport of CO2 and H(+) are described in Section 2. Since the rest of the chapter is dependent on these key mechanisms, they are given in detail, including the role of Jacobs-Stewart Cycle and its interaction with carbonic anhydrase. Also, this section deals briefly with the mechanisms of membrane depolarization of the chemoreceptor cells because this is one mechanism on which the responses depend. The metabolic impact of endogenous CO2 appears in the section with a historical twist, in the context of acclimatization to high altitude (Section 3). Because low P(O2) at high altitude stimulates the peripheral chemoreceptors (PC) increasing ventilation, the endogenous CO2 is blown off, making the internal milieu alkaline. With acclimatization however ventilation increases. This alkalinity is compensated in the course of time by the kidney and the acidity tends to be restored, but the acidification is not great enough to increase ventilation further. The question is what drives ventilation during acclimatization when the central pH is alkaline? The peripheral chemoreceptor came to the rescue. Its sensitivity to P(O2) is increased which continues to drive ventilation further during acclimatization at high altitude even when pH is alkaline. This link of CO2 through the O2 chemoreceptor is described in Section 4 which led to hypoxia-inducible factor (HIF-1). HIF-1 is stabilized during hypoxia, including the carotid body (CB) and brain cells, the seat of CO2 chemoreception. The cells are always hypoxic even at sea level. But how CO2 can affect the HIF-1 in the brain is considered in this section.
CO2 sensing in the central chemoreceptors (CC) is given in Section 5. CO(2)/H(+) is sensed by the various structures in the central nervous system but its respiratory and cardiovascular responses are restricted only to some areas. How the membranes are depolarized by CO2 or how it works through Na(+)/Ca(2+) exchange are discussed in this section. It is obvious, however, that CO2 is not maintained constant, decreasing with altitude as alveolar P(O2) decreases and ventilation increases. Rather, it is the [H(+)] that the organism strives to maintain at the expense of CO2. But then again, [H(+)] where? Perhaps it is in the intracellular environment.
Gap junctions in the carotid body and in the brain are ubiquitous. What functions they perform have been considered in Section 6. CO2 changes take place in lung alveoli where inspired air mixes with the CO2 from the returning venous blood. It is the interface between the inspired and expired air in the lungs where CO2 change is most dramatic. As a result, various investigators have looked for CO2 receptors in the lung, but none have been found in the mammals. Instead, CO2/H(+) receptors were found in birds and amphibians. However, they are inhibited by increasing CO2/H(+), instead of stimulated. But the afferent impulses transmitted to the brain produced stimulation in the efferents. This reversal of afferent-efferent inputs is a curious situation in nature, and this is considered in Section 7.
The NO and CO effects on CO2 sensing are interesting and have been briefly mentioned in Section 8.
A model for CO2/H(+) sensing by cells, neurons and bare nerve endings are also considered. These NO effects, models for CO2/H(+) and O2-sensitive cells in the CNS have been considered in the perspectives. Finally, in conclusion, the general theme of constancy of internal environment for CO2/H(+) is reiterated, and for that CO2/H(+) sensors-receptors systems are essential.
Since CO2/H(+) sensing as such has not been reviewed before, the recent findings in addition to defining basic CO2/H(+) reactions in the cells have been briefly summarized.