Bode bilder her som viser nevral tensjon fra Briegs arbeid på 70-tallet.
Bode bilder her som viser nevral tensjon fra Briegs arbeid på 70-tallet.
Nevner hvordan stretch øvelser av brystkassen sammen med innpust gir en lettere og roligere pust etterpå, og større bevegelse av diafragma. Dette prinsippet brukes i øvelsene i vårt Breathing System Diafragma trening.
Does Intercostal Stretch Alter Breathing Pattern and Respiratory Muscle Activity in Conscious Adults?
http://www.sciencedirect.com/science/article/pii/S0031940605609327
The effects of intercostal (IC) stretch on breathing patterns and respiratory muscle activity were monitored in nine healthy subjects. Tidal volume (Vt), breathing frequency (Fb), and inspiratory (Ti) and expiratory (Te) durations were determined from a pneumotachometer. Peak amplitudes and burst durations of activity in the diaphragm, parasternal ICs and external abdominal oblique muscles were determined from surface EMGs.
The third and eighth IC spaces were stretched in phase with inspiration or expiration when supine and 60° semi recumbent. Vt increased and Ti and Te were prolonged, resulting in a decreased Fb, independent of site of stretch, phase of breathing, or body position, during IC stretch compared to controls.
Peak amplitudes and burst durations of diaphragmatic EMG and burst durations of parasternal ICs were greater when the third and eighth IC spaces were stretched during inspiration compared to controls. Peak amplitudes of parasternal ICs increased only when the third IC space was stretched during inspiration. When applied during expiration, IC stretch increased only parasternal activity in the supine position. Intercostal stretch applied in phase with inspiration resulted in a slower, deeper breathing pattern with increased activity of the diaphragm and parasternal IC muscles. IC stretch may alter breathing sufficiently to improve gas exchange in some patients with pulmonary disorders.
EFFECT OF INTERCOSTAL STRETCH ON PULMONARY FUNCTION PARAMETERS AMONG HEALTHY MALES
Klikk for å få tilgang til Mohan_15062012_proof.pdf
The use of manual stretching procedures has become more prevalent in cardiorespiratory physiotherapy to improve pulmonary functions. However, limited evidence exists regarding evaluation of their effectiveness. The study aimed to determine the impact of Intercostal (IC) stretch in improving the dynamic pulmonary function parameters (Forced Expiratory Volume in the first second (FEV1), Forced Vital Capacity (FVC) and FEV1/FVC % and respiratory rate among healthy adults. Thirty healthy male subjects were recruited based on inclusion and exclusion criteria. Subjects were assigned to the experimental group and the control group through random sampling method. In the experimental group, subjects underwent IC stretch for ten breaths on the inspiratory phase of the respiratory cycle with breathing control exercises in semi recumbent position, while in the control group, breathing control exercises alone were performed in the semi recumbent position. The results of the study showed, FEV1/FVC % in the experimental group significantly improved with P=0.017 (p<0.05) than the control group, which means IC stretch increased lung volume and lead to improved lung function. This study suggested the IC stretching with breathing control may be more effective in improving dynamic lung parameters especially FEV1/FVC % than breathing control alone.
Nevner det aller meste om perifere nerver og hvordan de får plager av «normale» sitasjoner, som f.eks. musearm.
http://www.ncbi.nlm.nih.gov/pubmed/16386065
http://ptjournal.apta.org/content/86/1/92.long
The structural organization of peripheral nerves enables them to function while tolerating and adapting to stresses placed upon them by postures and movements of the trunk, head, and limbs. They are exposed to combinations of tensile, shear, and compressive stresses that result in nerve excursion, strain, and transverse contraction. The purpose of this appraisal is to review the structural and biomechanical modifications seen in peripheral nerves exposed to various levels of physical stress. We have followed the primary tenet of the Physical Stress Theory presented by Mueller and Maluf (2002), specifically, that the level of physical stress placed upon biological tissue determines the adaptive response of the tissue. A thorough understanding of the biomechanical properties of normal and injured nerves and the stresses placed upon them in daily activities will help guide physical therapists in making diagnoses and decisions regarding interventions.

Figure 1.
Structural components of peripheral nerves. In the endoneurial compartment (En), a single Schwann cell envelops several unmyelinated axons, and another Schwann cell provides multiple wrappings of plasma membrane forming the myelin sheath of a myelinated axon. The portion of a myelinated axon myelinated by a single Schwann cell is called the internode, and internodes are separated by nodes of Ranvier. Schwann cells associated with both unmyelinated and myelinated axons are covered with a continuous basal lamina (BL). Capillaries (Cap) are present within the endoneurial compartment, and collagen fibers (Col) run primarily longitudinally between the axons. The axons, Schwann cells, collagen, and endoneurial fluid are bundled into a fascicle by the perineurium (Pe). The perineurium consists of several layers of flattened perineurial cells connected by tight junctions and covered internally and externally by a basal lamina. The layers of perineurial cells are separated by collagen fibers (Col) oriented obliquely. Several fascicles are bundled together by the epineurium (Ep) to form a nerve. The epineurium consists primarily of fibroblasts, collagen fibers (Col), and elastic fibers. The epineurium between fascicles is termed the interfascicular epineurium, and that encompassing all of the fascicles is termed the epifascicular epineurium. Arterioles (A) and veins are oriented primarily longitudinally within the epineurium.
Blood supply
The blood supply to nerves is provided by coiled segmental arteries that enter the epineurium periodically along the length of the nerve and form the vasa nervorum. Arteries divide into epineurial arterioles that form an anastomotic network running primarily longitudinally within the epifascicular epineurium and the interfascicular epineurium (Fig. 3). Epineurial arterioles are supplied with a perivascular plexus of serotoninergic, adrenergic, and peptidergic nerves.17,18 Perforating arterioles cross the perineurium at oblique angles and carry a short sleeve of perineurial cells into the fascicle.3,19 Perineurial arterioles have poorly developed smooth muscle and thus have limited ability to regulate intrafascicular blood flow.20 Within the endoneurium, arterioles immediately turn into large-diameter, longitudinally oriented capillaries that allow blood flow in either direction (Fig. 3).21 The endothelial cells of endoneurial capillaries are connected by tight junctions, thus forming the tight blood-nerve barrier.7 Venules return blood to the venous system. Of note, lymphatic capillaries are present only within the epineurium; there is no lymphatic drainage from the intrafascicular or endoneurial space.22
Biomechanical properties
Under normal physiological conditions imposed by posture and movement, nerves are exposed to various mechanical stresses. Stress is defined as force divided by the area over which it acts9,23–25 and can be applied to a nerve as tensile, compressive, or shear stress or as a combination of stresses (Fig. 4). Tensile stress may be applied to tissues either parallel or perpendicular to the length of the nerve, causing respective longitudinal or transverse stress in the nerve. When joint motion causes elongation of the nerve bed, the nerve is inherently placed under tensile stress and accommodates the stress by both elongating and gliding.15 The deformation or change in nerve length induced by longitudinal tensile stress is called strain and is expressed typically as percent elongation.23,26–28 Displacement or gliding of a nerve relative to the surrounding nerve bed is called excursion.29–31 The direction of excursion may be longitudinal or transverse, or both, relative to the nerve tract31,32 and is measured in millimeters.

Figure 4.
Physical stresses placed on peripheral nerve. Tensile stress applied longitudinally to peripheral nerve creates an elongation of the nerve (an increase in strain). The transverse contraction that occurs during this elongation is greatest at the middle of the section undergoing tensile stress.
When the nerve bed is elongated, the nerve is placed under increased tensile stress. With the elongation of the nerve bed, the nerve glides toward the moving joint,1,33,34 a movement termed convergence.1 Conversely, if the nerve bed tension is relieved during joint motion, the nerve will realign along the shortened nerve bed by gliding away from the moving joint, a movement termed divergence.33 Convergence in the median nerve may be demonstrated during elbow extension (Fig. 5). The motion elongates the bed of the median nerve, causing the nerve segment in the arm to glide distally toward the elbow and the nerve segment in the forearm to glide proxi mally toward the elbow. In contrast, elbow extension relieves the tensile stresses in the ulnar nerve bed, causing the ulnar nerve to diverge away from the elbow (Fig. 5).

Figure 5.
Excursion of the median nerve (solid line) and the ulnar nerve (dotted line) during elbow extension followed by wrist extension. The concepts of nerve convergence toward and divergence away from a moving joint are illustrated in measurements of excursion taken at each site indicated. All measurements are reported in millimeters of proximal (P) or distal (D) excursion. The direction of excursion is also represented by solid arrows for median nerve excursion and open arrows for ulnar nerve excursion. (A) With elbow extension from 90° of flexion to 0° of flexion, the median nerve bed lengthens and the median nerve glides toward the elbow (converges). With the same joint motion, the ulnar nerve bed shortens and the ulnar nerve glides away from the elbow (diverges). (B) With wrist extension from 0° of extension to 60° of extension, both nerve beds lengthen; thus, both nerves converge toward the wrist. The magnitude of excursion is greatest closest to the moving joint. Data were obtained from: aDilley et al,29 bWright et al,27 and cWright et al.33 Measurements of nerve excursion at the wrist and elbow in panel A were extrapolated from studies of nerve excursion during elbow flexion from 0° to 90°.27
Nerve Stiffness
First, a recent study43 measured greater nerve compliance in nerve segments that cross joints than in segments that do not cross joints.
Second, nerve stiffness is greater in long nerve sections and in nerve sections with numerous branches.15 Severing nerve branches or vessels but leaving the nerve in situ results in increased compliance and decreased stiffness.15
Third, nerve stiffness is greater when a nerve is elongated rapidly rather than slowly. In addition, the ultimate strain at the point of failure appears to be dependent on the rate of elongation.
When a nerve is placed under tension and maintained at that new fixed length over time, there is a reduction in the tension in the nerve or the force required to maintain the fixed length. The observed reduction in tension may be plotted in a stress-relaxation curve (Fig.8).25,44 The majority of relaxation occurs in the first 20 minutes of fixed elongation.25,44Stress relaxation in nerves that are stretched slowly is greater than in nerves that are stretched rapidly.25,37,44–46 This phenomenon was observed when comparisons were made for rabbit tibial nerves stretched at different rates to lengths 6% longer than their resting lengths. Over the 60-minute relaxation time, there was a 57% reduction in stress in nerves elongated at 0.08% per second,45 but only a 34% reduction in stress in nerves elongated at 3.0% per second.44

Figure 8.
Stress-relaxation curve demonstrating viscoelastic properties of peripheral nerve. When a nerve is elongated and the new length is kept constant, there is a rapid reduction in the stress within the nerve, expressed as percent reduced relaxation. Most of the relaxation occurs in the first 20 minutes. The degree of elongation affects the amount of stress relaxation that will occur. The dotted line represents a nerve that has been elongated to 6% above its resting length. The solid line represents nerves that have been elongated to 9% and 12% above their resting lengths. Greater stress relaxation was documented in nerves that underwent less elongation.25,44 Modified from Kwan MK, Wall EJ, Massie J, Garfin SR. Strain, stress, and stretch of peripheral nerve: rabbit experiments in vitro and in vivo. Acta Orthop Scand. 1992;63:267–272, with permission of Taylor and Francis AS.
However, a nerve stretched repetitively to 8% or 10% strain exhibits a reduced slope of the stress-strain curve, indicating that that nerve undergoes less stress with successive elongations because of increased compliance and decreased stiffness.
Compression of nerve
In addition to tensile stress, nerves are exposed statically and dynamically to compressive stresses. As mentioned previously, the laws of physics dictate that the cross-sectional area of a cylindrical object is reduced as the cylinder is elongated. As a nerve is elongated under tensile force, the nerve undergoes transverse contraction, which is resisted by the fluid and nerve tissue contained within the connective tissue sheath.15,39The magnitude of the transverse contraction stress is greatest at the center of the elongating segment15 (Fig. 4). Nerves also may be compressed externally by approximation to adjacent tissues, such as muscle, tendon, or bone, or by pressure increases in the extraneural environment. Compression of a nerve segment causes displacement of its internal contents in transverse and longitudinal directions. As shown in rat nerve, extraneural compression causes an immediate displacement of endoneurial fluid to the edges of a compressive cuff over 5 to 10 minutes and a much slower displacement of axonal cytoplasm over the course of hours.48 The damage to axons and myelin is greatest at the edges of the compressed zone,48,49 where the shear forces are highest.50
At the edges of the cuff, however, myelin retraction with resultant widening of nodes and paranodal demyelination occurred. These structural alterations in myelin may be expected to result minimally in impaired impulse conduction or maximally in demyelination and a conduction block.
In response to biomechanical stresses placed on a nerve as an individual assumes a posture or movement, the nerve follows the path of least resistance.29 Combinations of tensile, shear, and compressive stresses result in combinations of nerve excursion, strain, and transverse contraction. Because the biomechanical forces on the nerve are so intricately linked, the sequencing and range of joint movement affect the magnitude and direction of excursion,27,29 the magnitude of nerve strain,27,29,35 and the degree of transverse contraction at different sites along the nerve.27
Simultaneous nerve excursion, strain, and transverse contraction may be seen in the ulnar nerve as an example of responses to physical stresses imposed during movements of the upper limb. When the upper limb is maintained in a position of 90 degrees of shoulder abduction and 90 degrees of shoulder external rotation with the wrist neutral, and when the elbow is moved from 90 degrees of flexion to full extension, the ulnar nerve bed is shortened and the tensile stress on the nerve is decreased. With this motion, there is divergence of the ulnar nerve away from the elbow (Fig. 5), decreased nerve strain, especially at the elbow (Fig. 6), and decreased compression within the cubital tunnel.27,29,33 When the wrist then is extended from neutral to full extension, the ulnar nerve bed is lengthened, resulting in convergence of the nerve toward the wrist (Fig. 5), an increase in nerve strain (Fig. 6), and transverse contraction greatest in the nerve segment across the carpal bones and at the tunnel of Guyon.27,29,33 The magnitude of nerve strain and excursion will be greatest near the wrist, and the fascicles will rearrange as the nerve assumes a flattened oval shape. Because the nerve does not lie directly on the rotational axis of joint motion, the fascicles farthest from the axis will undergo greater strain than those closer to the center of rotation51

Figure 6.
Strain of the median nerve (solid line) and the ulnar nerve (dotted line) during elbow extension followed by wrist extension. Measurements at the sites indicated are reported as percent increase (↑) or percent decrease (↓) in strain. (A) With elbow extension from 90° of flexion to 0° of flexion, median nerve strain increases because of elongation of the nerve bed. Conversely, ulnar nerve strain decreases as the ulnar nerve bed shortens. (B) With wrist extension from 0° of extension to 60° of extension, the strain at the sites measured increases in both nerves as both nerve beds elongate. The magnitude of the strain is greatest closest to the moving joint. Data were obtained from: aWright et al27and bWright et al.33 Measurements of nerve excursion at the wrist and elbow in panel A were extrapolated from studies of nerve excursion during elbow flexion from 0° to 90°.27
Continuum of physical stress states
First, levels of physical stress lower than the levels required for tissue maintenance (low stress) result in a reduced ability of the tissue to tolerate subsequent stress and are consistent with tissue plasticity and response to functional demand.
Second, levels of physical stress in the range required for tissue maintenance (normal stress) result in no tissue adaptations and are considered to maintain a state of equilibrium.
Third, physical stress levels that exceed the range required for tissue maintenance (high stress) result in an increase in the tolerance of the tissue for stress in an effort to meet the mechanical demand.
Fourth, physical stress levels that exceed the capacity of some components of the tissue (excessive stress) result in tissue injury.
Fifth, levels of physical stress that are extreme (extreme stress) result in tissue death.
Finally, it is important to note that the physical stress level is a composite value with variable components of magnitude, time, and direction or type of stress.
In the functional zone, the physical stresses on the nerve are sufficient to maintain a state of equilibrium and normal physiological function. In the dysfunctional zone, various levels of physical stress have altered the ability of the nerve to tolerate subsequent stress.

Figure 9.
Continuum of physical stress states. The white area represents the functional zone in which the physical stresses on the nerve are sufficient to maintain a state of equilibrium and normal physiological function. The shaded areas represent dysfunctional zones resulting from various levels of physical stress placed on the nerve tissue. Under conditions of prolonged low stress, the functional zone will shrink in width and shift to the left, reducing the ability of the tissue to tolerate subsequent stresses even of previously normal levels. Under conditions of high stress, the functional zone may expand and shift to the right, improving the ability of the tissue to tolerate subsequent physical stress. If the nerve is exposed to prolonged or repeated excessive stress, the functional zone will shrink in width. Although scarring of damaged tissue may enable the nerve to tolerate subsequent physical stresses, the physiological function of the nerve will be reduced. Exposure to extreme stress will result in disruption of axon continuity or neural cell death and significantly reduced physiological function.
Under conditions of immobilization, such as casting, splinting, and bracing, peripheral nerves are exposed to levels of physical stress that are lower than those necessary to maintain the nerves in a state of equilibrium or in a functional zone (Fig. 9). According to the Physical Stress Theory, nerve will undergo predictable physiological and structural modifications proportional to the levels of reduced stress and the duration of immobilization.2 Immobilization induces cell biological changes in axons and axon terminals52–54 and structural changes in myelin and nerve connective tissue layers that likely alter the ability of nerves to tolerate subsequent physical stress.
We hypothesize that after a period of immobilization, the width of the functional zone on the continuum of physical stress states will shrink and shift toward the left (Fig. 9).
Nerves are exposed to various levels of longitudinal tensile stress during limb-lengthening procedures, such as distraction osteogenesis (Ilizarov procedures), traction injuries, and stretching maneuvers. The tissue response is dependent upon the magnitude and duration of the tensile stress. The extant data indicate that lengthening of 6% to 8% for a short duration causes transient physiological changes that appear to be within the normal stress tolerance of the tissue, whereas acute strains of 11% and greater cause long-term damage and may be considered to be excessive or extreme stress states.
In cadavers, positioning in shoulder depression, 90 degrees of shoulder abduction, 90 degrees of shoulder external rotation, 70 degrees of forearm supination, 60 degrees of wrist extension, full finger extension, and full elbow extension resulted in 7.6%±8.2% (X̄±SD) strain in the median nerve measured just proximal to the wrist.28 Adults who were healthy and who were placed in this same position lacked 12±13 degrees (X̄±SD) of elbow extension because of substantial discomfort in the limb.61 The subjects reported pain of 5.1±1.9 (X̄±SD) on a 10-point visual analog scale, and 36% of the subjects reported paresthesia in the upper limb. Taken together, these findings suggest that many people are unable to tolerate levels of strain below the theoretical 11% threshold.
Compression on a nerve may be the result of extraneural force or may occur as transverse contraction secondary to increased longitudinal strain (Fig. 4). Compression stress of a low magnitude and a short duration may result in reversible physiological and minor structural changes. Compressive stress of a high magnitude, however, may result in structural alterations in myelin sheaths and even disruption of axons. Low-magnitude compressive stress applied over a long period of time may cause significant structural changes in the nerve secondary to impairment of blood flow and sequelae of ischemia.
As with strain-induced injury, a threshold for compression-induced nerve injury is difficult to determine. Common functional positions may result in compression pressures that approach or exceed the 20 to 30 mm Hg demonstrated to impair nerve blood flow.75 The carpal tunnel is a site well known for compressive damage to the median nerve and thus has been well studied. Carpal tunnel pressure in subjects who were healthy was measured at 3 to 5 mm Hg with the wrist in a neutral position.76–78 Simply placing the hand on a computer mouse was shown to increase the tunnel pressure from the resting 5 mm Hg to 16 to 21 mm Hg,79 and actively using the mouse to point and click increased the tunnel pressure to 28 to 33 mm Hg, a pressure high enough to reduce nerve blood flow.
In subjects with carpal tunnel syndrome, pressure in the carpal tunnel was 32 mm Hg with the wrist in a neutral position and rose to a mean of 110 mm Hg with full wrist extension in subjects with carpal tunnel syndrome.76 These tunnel pressures exceed the threshold of 20 to 30 mm Hg for vascular perfusion even at rest. Taken together, these findings suggest that even functional positions, such as the use of a computer keyboard and mouse, place the wrist in a position of increased carpal tunnel pressure, compromising nerve blood flow and placing people at risk for median nerve injury.
Direct damage to myelin and axons has been shown to occur with extraneural compression of as low as 50 mm Hg maintained for 2 minutes,48 and the percentage of damaged fibers increases with increasing force. Ten days after the application of compressive stress at 50 mm Hg, 30% of the axons in the region under the compressive cuff showed evidence of demyelination, focal myelin thickening, remyelination, and axonal degeneration or regeneration.48
The pathological consequences of prolonged compression include subperineurial edema; inflammation; deposition of fibrin; activation of endoneurial fibroblasts, mast cells, and macrophages; demyelination; axon degeneration; and fibrosis.83 Compression of a very long duration has been modeled in animals with loose ligatures,88 Silastic* tubes,89,90and pressure balloons placed within an anatomical tunnel.91 The pathological findings are thought to result from both inflammatory and cellular phenomena and include changes in the blood-nerve barrier, thickening of the perineurium and epineurium, thinning of myelin, demyelination and degeneration of axons in the fascicle periphery, and slowed nerve conduction velocity.
In the case of chronic compression, decompression is paramount. Physical therapy intervention should focus on reduction of inflammation, improvement in blood flow, and enhancement of the capacity of the nerve for strain and excursion along its full length in an effort to reduce the physical stress on the compressed region.
Vibration constitutes one form of repetitive stress. We know from studies of humans who use hand-held vibrating tools that vibration stresses can cause reductions in tactile sensation, as well as other sensory disturbances96 and reduced grip force.97,98Furthermore, myelin breakdown and fibrosis have been seen in the dorsal interosseous nerve at the wrist in people with vibration-induced neuropathy.99 Long-term exposure to vibration stresses has been shown to result in the grouping of muscle fiber types in muscle biopsies, indicative of denervation and reinnervation.98
Repetitive movements, such as those that occur in work-related musculoskeletal disorders, were discussed in detail recently by Barr and Barbe.102 The stresses placed upon the tissues may be variable in terms of type, magnitude, frequency, and duration, and the combination of these factors may place nerves in normal to extreme levels of physical stress. The chronic inflammation associated with repetitive movements places nerves under constantly higher hydrostatic compressive stress, which may increase further with contraction of the surrounding muscles. Chronic inflammation elicits within the nerves a remodeling response that seeks to add mechanical stability.103 The most common outcome is the deposition of collagen in the connective tissue layers, which leads to decreased compliance of the nerves to elongation. As with chronic compression, the approach for assessment and treatment of injuries attributable to repetitive movements must address the chronic inflammatory state and connective tissue changes. Of primary importance in interventions for all stress-induced injuries are the identification and characterization of physical stresses and the modification of their components, magnitude, time, and direction, as outlined in the physical stress theory.2
Summary
This assessment should guide treatment interventions to normalize the stresses on the nerves, be they rest, soft tissue or neurodynamic mobilization, stretching, modalities, exercise, or patient education. Treatment rationale should be based on an educated understanding of the biomechanical properties of normal and pathological nerves. The concept of a continuum of low-normal-high-excessive-extreme stresses may be used as a training tool for patient education, pointing out examples of daily activities that fall under the different categories.
Alt om smerte, kort fortalt, fra: http://www.somasimple.com/forums/showthread.php?t=4942
Nothing Simple – Ten Steps to Understanding Manual and Movement Therapies for Pain
1. Pain is a category of complex experiences, not a single sensation produced by a single stimulus.
2. Nociception (warning signals from body tissues) is neither necessary nor sufficient to produce pain. In other words, pain can occur in the absence of tissue damage.
3. A pain experience may be induced or amplified by both actual and potential threats.
4. A pain experience may involve a composite of sensory, motor, autonomic, endocrine, immune, cognitive, affective and behavioural components. Context and meaning are paramount in determining the eventual output response.
5. The brain maps peripheral and central neural processing into each of these components at multiple levels. Therapeutic input at a single level may be sufficient to resolve a threat response.
6. Manual and movement therapies may affect peripheral and central neural processes at various stages:
– transduction of nociception at peripheral sensory receptors
– transmission of nociception in the peripheral nervous system
– transmission of nociception in the central nervous system
– processing and modulation in the brain
7. Therapies that are most likely to be successful are those that address unhelpful cognitions and fear concerning the meaning of pain, introduce movement in a non-threatening internal and external context, and/or convince the brain that the threat has been resolved.
8. The corrective physiological mechanisms responsible for resolution are inherent. A therapist need only provide an appropriate environment for their expression.
9. Tissue length, form or symmetry are poor predictors of pain. The forces applied during common manual treatments for pain generally lack the necessary magnitude and specificity to achieve enduring changes in tissue length, form or symmetry. Where such mechanical effects are possible, the clinical relevance to pain is yet to be established. The predominant effects of manual therapy may be more plausibly regarded as the result of reflexive neurophysiological responses.
10. Conditioning for the purposes of fitness and function or to promote general circulation or exercise-induced analgesia can be performed concurrently but points 6 and 9 above should remain salient.
Bibliography
Books:
Pain: The Science of Suffering – Patrick Wall
The Challenge of Pain – Patrick Wall, Ronald Melzack
Explain Pain – David Butler, Lorimer Moseley
The Sensitive Nervous System – David Butler
Phantoms in the Brain – V. S. Ramachandran
Topical Issues in Pain Vol’s 1-5 – Louis Giffiord (ed)
The Feeling of What Happens – Antonio Damasio
Clinical Neurodynamics – Michael Shacklock
Eyal Lederman – The Science and Practice of Manual Therapy
Research articles:
Melzack R. Pain and the neuromatrix in the brain. J Dental Ed. 2001;65:1378-82.
Craig AD. Pain mechanisms: Labeled lines versus convergence in central processing. Ann Rev Neurosci. 2003;26:130.
Craig AD. How do you feel? Interoception: the sense of the physiological condition of the body. Nature Rev Neurosci. 2002;3:655-66.
Henderson LA, Gandevia SC, Macefield VG. Somatotopic organization of the processing of muscle and cutaneous pain in the left and right insula cortex: A single-trial fMRI study. Pain. 2007;128:20-30.
Olausson H, Lamarre Y, Backlund H, Morin C, Wallin BG, Starck G, Ekholm S, Strigo I, Worsley K, Vallbo AB, Bushnell MC. Unmyelinated tactile afferents signal touch and project to insular cortex. Nature Neurosci. 2002;5:900–904.
Moseley GL. A pain neuromatrix approach to patients with chronic pain. Manual Ther. 2003;8:130-40.
Moseley GL. Unravelling the barriers to reconceptualisation of the problem in chronic pain: The actual and perceived ability of patients and health professionals to understand the neurophysiology. J Pain. 2003;4:184-89.
Moseley GL, Arntz A. The context of a noxious stimulus affects the pain it evokes. Pain. 2007;133(1-3):64-71.
Moseley, GL, Nicholas, MK and Hodges, PW. A randomized controlled trial of intensive neurophysiology education in chronic low back pain. Clin J Pain. 2004;20:324-30.
Crombez G, Vlaeyen JWS, Heuts PH et al. Pain-related fear is more disabling than pain itself. Evidence on the role of pain-related fear in chronic back pain disability. Pain. 1999;80:329-40.
Zusman M. Forebrain-mediated sensitization of central pain pathways: ‘non-specific’ pain and a new image for manual therapy. Manual Ther. 2002;7:80-88.
Dorko B. The analgesia of movement: Ideomotor activity and manual care. J Osteopathic Med. 2003;6:93-95.
Threlkeld AJ. The effects of manual therapy on connective tissue. Phys Ther. 1992;72:893-902.
Lederman E. The myth of core stability. Retrieved at: http://www.ppaonline.co.uk/
Svært interessant studie som viser at å strekke bindevev jevnlig, f.eks. slik vi gjør under behandling eller i yoga, gjør at vi får mindre arrvev. Det produseres mindre TGF-B1, et molekyl som stimulerer arrvevproduksjon. Spesielt for indre organer er dette hjelpsomt, som lunger og tarmer, f.eks. etter operasjoner eller ved betennelsesykdommer. Forskerene viser at det holder å strekke overkroppen så det blir 20-30% lengre avstand mellom hofte og skulder. Dette får vi til med noe så enkelt som å svaie ryggen og strekke armene opp. F.eks. ved å ligge på ryggen over en ball eller gjøre The Founder.

http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3065715/
We tested the hypothesis that brief (10 min) static tissue stretch attenuates TGF-β1-mediated new collagen deposition in response to injury.
In the in vivo model, microinjury resulted in a significant increase in Type-1 procollagen in the absence of stretch (P < 0.001), but not in the presence of stretch (P = 0.21). Thus, brief tissue stretch attenuated the increase in both soluble TGF-β1 (ex vivo) and Type-1 procollagen (in vivo) following tissue injury. These results have potential relevance to the mechanisms of treatments applying brief mechanical stretch to tissues (e.g., physical therapy, respiratory therapy, mechanical ventilation, massage, yoga, acupuncture).
Transforming growth factor β1 (TGF-β1) is well-established as one of the key cytokines regulating the response of fibroblasts to injury, as well as the pathological production of fibrosis (Barnard et al., 1990;Sporn and Roberts, 1990; Leask and Abraham, 2004). Tissue injury is known to cause auto-induction of TGF-β1 protein production and secretion (Van Obberghen-Schilling et al., 1988; Morgan et al., 2000). Elevated extracellular levels of TGF-β1 have a major impact on extracellular matrix composition by causing autocrine and paracrine activation of fibroblast cell surface receptors, leading to increased synthesis of collagens, elastin, proteoglycans, fibronectin, and tenascin (Balza et al., 1988; Bassols and Massague, 1988; Kahari et al., 1992; Cutroneo, 2003).
In vivo, connective tissue remodeling is not limited to tissue injury, but also occurs in response to changing levels of tissue mechanical forces (e.g., immobilization, beginning a new exercise or occupation). Long-standing physical therapy practices also suggest that externally applied mechanical forces can be used to reduce collagen deposition during tissue repair and scar formation (Cummings and Tillman, 1992).
In the stretch group, the mice underwent stretching of the trunk for 10 min twice a day for 7 days in the following manner: each mouse was suspended by the tail such that its paws barely touched a surface slightly inclined relative to the vertical. In response to this maneuver, the mouse spontaneously extended its front and hind limbs (Fig. 1B) with the distance between ipsilateral hip and shoulder joints becoming 20–30% greater than the resting distance.
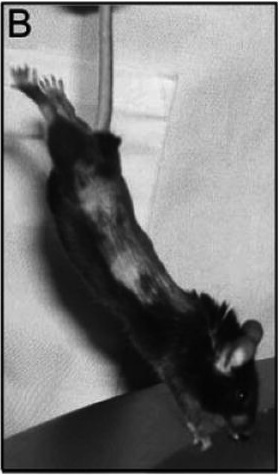
B: Method used to induce tissue stretch in vivo. Mice are suspended by the tail such that their paws barely touch a surface slightly inclined relative to the vertical. The mice spontaneously extend their front and hind limbs, the distance between ipsilateral hip and shoulder joints becoming 20–30% greater than the resting distance.

Effect of tissue stretch on TGF-β1 protein ex vivo. A: Time course of TGF-β1 protein levels in the culture media for non-stretched (closed circle, N = 4) and stretched (open circle, N = 4) mouse subcutaneous tissue explants on days 0, 1, and 3 post-stretch (or no stretch). All tissue samples were excised and incubated for 24h prior to day 0.B:Levels of TGF-β1 protein in the culture media at day 3 for non-stretched and stretched sbcutaneous tissue samples (N = 36). Asterisk (*) indicates significant difference from stretched (P = 0.002). Error bars represent standard errors.

Ex vivo tissue injury and cell viability assessment. A: Time course of LDH concentration in the culture media (marker of cell death) for non-stretched (closed circle, n = 4) and stretched (open circle, n = 4) mouse subcutaneous tissue explants on days 0, 1, and 3 post-stretch (or no stretch). B,C: Confocal microscopy imaging of mouse subcutaneous tissue explants showing similar proportions of live (green) and dead (red) cells in non-stretched (A) versus stretched (B) tissue after 3 day incubation post-stretch (or no stretch). Images are projections of three-dimensional image stacks. Scale bars: 40 μm.

Effect of tissue stretch in vivo on subcutaneous tissue Type-1 procollagen in mouse microinjury model. A: Mean ± SE procollagen percent staining area in non-injured versus injured sides, without stretch (N = 11) and with stretch (N = 10); B,C: Type-1 procollagen in non-stretched and stretched tissue (both injured). Scale bars, 40 μm.
First, stretching mouse subcutaneous tissue explants by 20% for 10 min decreases soluble TGF-β1 levels measured 3 days after stretch. During the 4-day incubation, TGF-β1 levels in the culture media increase in both stretched and non-stretched samples; because some tissue trauma occurs at the time of excision, this progressive rise in TGF-β1 is consistent with an injury response. However, the increase in the level of TGF-β1 is slower in the samples that are briefly stretched for 10 min, compared with samples that are not stretched. Since TGF-β1 auto-induction is an important mechanism driving the increase in collagen synthesis following tissue injury (Cutroneo, 2003), we hypothesized that brief stretching of tissue following injury in vivo would decrease soluble TGF-β1 levels, attenuate TGF-β1 auto-induction and decrease new collagen deposition.
Testing this hypothesis in a mouse subcutaneous tissue injury model showed that elongating the tissues of the trunk by 20–30% for 10 min twice a day significantly reduces the amount of subcutaneous new collagen 7 days following subcutaneous tissue injury.
Reducing scar and adhesion formation using stretch and mobilization is especially important for internal tissue injuries and inflammation involving fascia and organs, as opposed to open wounds. For open wounds (including surgical incisions) and severe internal tears (such as a ruptured ligament or tendon), wound closure and strength are critical and thus a certain amount of scarring is necessary and inevitable. In the case of minor sprains and repetitive motion injuries, however, scarring is mostly detrimental since it can contribute to maintaining the chronicity of tissue stiffness, abnormal movement patterns, and pain (Langevin and Sherman, 2007).
We have proposed that therapies that briefly stretch tissues beyond the habitual range of motion (physical therapy, massage, yoga, acupuncture) locally inhibit new collagen formation for several days after stretch and thus prevent and/or ameliorate soft tissue adhesions (Langevin et al., 2001, 2002, 2005, 2006a, 2007).

Proposed model for healing of connective tissue injury in the absence (A,C,E) and presence (B,D,F) of tissue stretch. In this model, brief stretching of tissue beyond the habitual range of motion reduces soluble TGF-β1 levels (D) causing a decrease in the fibrotic response, less collagen deposition, and reduced tissue adhesion (F) compared with no stretch (E). Black lines represent newly formed collagen.
Om hvordan bindevevet i huden beveger seg når man strekker huden. Nevner at nerver og blodårer har veldig svingete baner i huden, og at dette gjør at vi tåler mye strekk og bevegelse uten at disse strekkes eller ødelegges.
http://www.ncbi.nlm.nih.gov/pubmed/14527168/
The subcutaneous connective tissue was observed to be composed of multiple layers of thin collagen sheets containing elastic fibers. These piled-up collagen sheets were loosely interconnected with each other, while the outer and inner sheets were respectively anchored to the dermis and epimysium by elastic fibers. Collagen fibers in each sheet were variable in diameter and oriented in different directions to form a thin, loose meshwork under conditions without mechanical stretching.
When a weak shear force was loaded between the skin and the underlying abdominal muscles, each collagen sheet slid considerably, resulting in a stretching of the elastic fibers which anchor these sheets. When a further shear force was loaded, collagen fibers in each sheet seemed to align in a more parallel manner to the direction of the tension. With the reduction or removal of the force, the arrangement of collagen fibers in each sheet was reversed and the collagen sheets returned to their original shapes and positions, probably with the stabilizing effect of elastic fibers.
Blood vessels and nerves in the subcutaneous connective tissue ran in tortuous routes in planes parallel to the unloaded skin, which seemed very adaptable for the movement of collagen sheets. These findings indicate that the subcutaneous connective tissue is extensively mobile due to the presence of multilayered collagen sheets which are maintained by elastic fibers.
Never det meste rundt bindevev: tensegritet, subcutan hud, skinligaments, stretching, ligamenter, nerver, m.m.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2667913/
Fasciae probably hold many of the keys for understanding muscle action and musculoskeletal pain, and maybe of pivotal importance in understanding the basis of acupuncture and a wide range of alternative therapies (Langevin et al. 2001, 2002, 2006a; Langevin & Yandow, 2002; Iatridis et al. 2003). Intriguingly, Langevin et al. (2007) have shown that subtle differences in the way that acupuncture needles are manipulated can change how the cells in fascia respond. The continuum of connective tissue throughout the body, the mechanical role of fascia and the ability of fibroblasts to communicate with each other via gap junctions, mean that fascia is likely to serve as a body-wide mechanosensitive signaling system with an integrating function analogous to that of the nervous system (Langevin et al. 2004; Langevin, 2006). It is indeed a key component of a tensegrity system that operates at various levels throughout the body and which has been considered in detail by Lindsay (2008) in the context of fascia.
Anatomists have long distinguished between superficial and deep fascia (Fig. 1), although to many surgeons, ‘fascia’ is simply ‘deep fascia’. The superficial fascia is traditionally regarded as a layer of areolar connective or adipose tissue immediately beneath the skin, whereas deep fascia is a tougher, dense connective tissue continuous with it.

A diagrammatic representation of a transverse section through the upper part of the leg showing the relative positions of the superficial (SF) and deep fascia (DF) in relation to the skin (S) and muscles. Note how the deep fascia, in association with the bones [tibia (T) and fibula (F)] and intermuscular septa (IS) forms a series of osteofascial compartments housing the extensor, peroneal (PER) and flexor muscles. If pressure builds up within a compartment because of an acute or overuse injury, then the vascular supply to the muscles within it can be compromised and ischaemia results. ANT, anterior compartment; IM, interosseous membrane.
The presence of a significant layer of fat in the superficial fascia is a distinctive human trait (thepanniculus adiposus), compensating for the paucity of body hair. It thus plays an important role in heat insulation. In hairy mammals, the same fascia is typically an areolar tissue that allows the skin to be readily stripped from the underlying tissues (Le Gros Clark, 1945). Where fat is prominent in the superficial fascia (as in man), it may be organized into distinctive layers, or laminae (Johnston & Whillis, 1950), although Gardner et al. (1960) caution that these may sometimes be a characteristic of embalmed cadavers and not evident in the living person. Furthermore, Le Gros Clark (1945) also argues that fascial planes can be artefactually created by dissection. Conversely, however, some layers of deep fascia are more easily defined in fresh than in fixed cadavers (Lytle, 1979).
The superficial fascia conveys blood vessels and nerves to and from the skin and often promotes movement between the integument and underlying structures.
Skin mobility protects both the integument and the structures deep to it from physical damage. Mobility is promoted by multiple sheets of collagen fibres coupled with the presence of elastin (Kawamata et al. 2003). The relative independence of the collagen sheets from each other promotes skin sliding and further stretching is afforded by a re-alignment of collagen fibres within the lamellae. The skin is brought back to its original shape and position by elastic recoil when the deforming forces are removed. As Kawamata et al. (2003)point out, one of the consequences of the movement-promoting characteristics of the superficial fascia is that the blood vessels and nerves within it must run a tortuous route so that they can adapt to an altered position of the skin, relative to the deeper structures.
Although deep fascia elsewhere in the limbs is often not so tightly bound to the skin, nevertheless cutaneous ligaments extending from deep fascia to anchor the integument are much more widespread than generally recognized and serve to resist a wide variety of forces, including gravitational influences (Nash et al. 2004).
According to Bouffard et al. (2008), brief stretching decreases TGF-β1-mediated fibrillogenesis, which may be pertinent to the deployment of manual therapy techniques for reducing the risk of scarring/fibrosis after an injury. As Langevin et al. (2005) point out, such striking cell responses to mechanical load suggest changes in cell signaling, gene expression and cell-matrix adhesion.
In contrast, Schleip et al. (2007) have reported myofibroblasts in the rat lumbar fascia (a dense connective tissue). The cells can contract in vitro andSchleip et al. (2007) speculate that similar contractions in vivo may be strong enough to influence lower back mechanics. Although this is an intriguing suggestion that is worthy of further exploration, it should be noted that tendon cells immunolabel just as strongly for actin stress fibres as do fascial cells and this may be associated with tendon recovery from passive stretch (Ralphs et al. 2002). Finally, the reader should also note that true muscle fibres (both smooth and skeletal) can sometimes be found in fascia. Smooth muscle fibres form the dartos muscle in the superficial fascia of the scrotum and skeletal muscle fibres form the muscles of fascial expression in the superficial fascia of the head and neck.
Consequently, entheses are designed to reduce this stress concentration, and the anatomical adaptations for so doing are evident at the gross, histological and molecular levels. Thus many tendons and ligaments flare out at their attachment site to gain a wide grip on the bone and commonly have fascial expansions linking them with neighbouring structures. Perhaps the best known of these is the bicipital aponeurosis that extends from the tendon of the short head of biceps brachii to encircle the forearm flexor muscles and blend with the antebrachial deep fascia (Fig. 6). Eames et al. (2007) have suggested that this aponeurosis may stabilize the tendon of biceps brachii distally. In doing so, it reduces movement near the enthesis and thus stress concentration at that site.

The bicipital aponeurosis (BA) is a classic example of a fascial expansion which arises from a tendon (T) and dissipates some of the load away from its enthesis (E). It originates from that part of the tendon associated with the short head of biceps brachii (SHB) and blends with the deep fascia (DF) covering the muscles of the forearm. The presence of such an expansion at one end of the muscle only, means that the force transmitted through the proximal and distal tendons cannot be equal. LHB, long head of biceps brachii. Photograph courtesy of S. Milz and E. Kaiser.
Several reports suggest that fascia is richly innervated, and abundant free and encapsulated nerve endings (including Ruffini and Pacinian corpuscles) have been described at a number of sites, including the thoracolumbar fascia, the bicipital aponeurosis and various retinacula (Stilwell, 1957; Tanaka & Ito, 1977; Palmieri et al. 1986; Yahia et al. 1992; Sanchis-Alfonso & Rosello-Sastre, 2000; Stecco et al. 2007a).
Changes in innervation can occur pathologically in fascia, and Sanchis-Alfonso & Rosello-Sastre (2000) report the ingrowth of nociceptive fibres, immunoreactive to substance P, into the lateral knee retinaculum of patients with patello-femoral malignment problems.
Stecco et al. (2008) argue that the innervation of deep fascia should be considered in relation to its association with muscle. They point out, as others have as well (see below in ‘Functions of fascia’) that many muscles transfer their pull to fascial expansions as well as to tendons. By such means, parts of a particular fascia may be tensioned selectively so that a specific pattern of proprioceptors is activated.
It is worth noting therefore that Hagert et al. (2007) distinguish between ligaments at the wrist that are mechanically important yet poorly innervated, and ligaments with a key role in sensory perception that are richly innervated. There is a corresponding histological difference, with the sensory ligaments having more conspicuous loose connective tissue in their outer regions (in which the nerves are located). Comparable studies are not available for deep fascia, although Stecco et al. (2007a) report that the bicipital aponeurosis and the tendinous expansion of pectoralis major are both less heavily innervated than the fascia with which they fuse. Where nerves are abundant in ligaments, blood vessels are also prominent (Hagert et al. 2005). One would anticipate similar findings in deep fascia.
Some of the nerve fibres associated with fascia are adrenergic and likely to be involved in controlling local blood flow, but others may have a proprioceptive role. Curiously, however, Bednar et al. (1995)failed to find any nerve fibres in thoracolumbar fascia taken at surgery from patients with low back pain.
The unyielding character of the deep fascia enables it to serve as a means of containing and separating groups of muscles into relatively well-defined spaces called ‘compartments’.
One of the most influential anatomists of the 20th century, Professor Frederic Wood Jones, coined the term ‘ectoskeleton’ to capture the idea that fascia could serve as a significant site of muscle attachment – a ‘soft tissue skeleton’ complementing that created by the bones themselves (Wood Jones, 1944). It is clearly related to the modern-day concept of ‘myofascia’ that is popular with manual therapists and to the idea of myofascial force transmission within skeletal muscle, i.e. the view that force generated by skeletal muscle fibres is transmitted not only directly to the tendon, but also to connective tissue elements inside and outside the skeletal muscle itself (Huijing et al. 1998; Huijing, 1999).
One can even extend this idea to embrace the concept that agonists and antagonists are mechanically coupled via fascia (Huijing, 2007). Thus Huijing (2007) argues that forces generated within a prime mover may be exerted at the tendon of an antagonistic muscle and indeed that myofascial force transmission can occur between all muscles of a particular limb segment.
Wood Jones (1944) was particularly intrigued by the ectoskeletal function of fascia in the lower limb. He related this to man’s upright stance and thus to the importance of certain muscles gaining a generalized attachment to the lower limb when it is viewed as a whole weight-supporting column, rather than a series of levers promoting movement. He singled out gluteus maximus and tensor fascia latae as examples of muscles that attach predominantly to deep fascia rather than bone (Wood Jones, 1944).
They have argued that a common attachment to the thoracolumbar fascia means that the latter has an important role in integrating load transfer between different regions. In particular, Vleeming et al. (1995) have proposed that gluteus maximus and latissimus dorsi (two of the largest muscles of the body) contribute to co-ordinating the contralateral pendulum like motions of the upper and lower limbs that characterize running or swimming. They suggest that the muscles do so because of a shared attachment to the posterior layer of the thoracolumbar fascia. Others, too, have been attracted by the concept of muscle-integrating properties of fascia. Thus Barker et al. (2007) have argued for a mechanical link between transversus abdominis and movement in the segmental neutral zone of the back, via the thoracolumbar fascia. They feel that the existence of such fascial links gives an anatomical/biomechanical foundation to the practice in manual therapy of recommending exercises that provoke a submaximal contraction of transversus abdominis in the treatment of certain forms of low back pain.
An important function of deep fascia in the limbs is to act as a restraining envelope for muscles lying deep to them. When these muscles contract against a tough, thick and resistant fascia, the thin-walled veins and lymphatics within the muscles are squeezed and their unidirectional valves ensure that blood and lymph are directed towards the heart. Wood Jones (1944) contests that the importance of muscle pumping for venous and lymphatic return is one of the reasons why the deep fascia in the lower limb is generally more prominent than in the upper – because of the distance of the leg and foot below the heart.
In certain regions of the body, fascia has a protective function. Thus, the bicipital aponeurosis (lacertus fibrosus), a fascial expansion arising from the tendon of the short head of biceps brachii (Athwal et al. 2007), protects the underlying vessels. It also has mechanical influences on force transmission and stabilizes the tendon itself distally (Eames et al. 2007).
En gjennomgang av hvordan sitteposisjon og holdning påvirker kroppen.
http://www.dynamicchiropractic.ca/mpacms/dc_ca/article.php?id=56598
In order to assess the loads placed on a spine during various positions, Rohlmann, et al. (2011) looked at various seating positions.4 They found the implant force increased 48 percent for 15 degrees flexion and decreased 19 percent for 10 degrees extension of the trunk. Placing the hands on the thighs reduced the loads by 19 percent, on average, compared to having arms hanging at the sides.
Dreischarf, et al. (2010) also found that reduced spinal load during sitting can be achieved by supporting the upper body with the arms.5
A study by De Carvalho, et al. (2010) compared lumbar spine and pelvic posture between standing and sitting via radiologic investigation. Lumbar lordosis and sacral inclination decreased by 43 and 44 degrees, respectively.6 This shows that with respect to sitting posture, to goal should be to maintain or prevent a reduction of the lumbar lordosis.
One study found 40-percent higher cervical extensor activity in the slouched posture. More neutral sitting postures reduce the demand on the cervical extensor muscles.7 Education on maintaining a neutral sitting posture can offset the detrimental effects.
A study by Caneiro, et al. (2010) showed that slumped sitting was associated with greater head / neck flexion, and increased muscle activity of the cervical erector spinae.9 Adjustments to seat angle and lumbar roll can also significantly effect head and neck posture.
A study by Horton, et al. (2010) found that the degree of angulation of the backrest support of an office chair, plus the addition of a lumbar roll support, are the two most important seat factors that will benefit head and neck postural alignment.10
A study by Bullock, et al. (2005) looked at how sitting posture can affect range of motion and pain for those with shoulder impingement.11 An erect posture appeared to increase active shoulder flexion, although there was no difference in shoulder pain between an erect and slouched posture.
Finley, et al. (2003) found that an increased thoracic kyphosis from a slouched posture can significantly alter the kinematics of the scapula during humeral elevation.12
And Kebaetse, et al. (1999) found that a slouched posture is associated with a 16.2 percent reduction in arm horizontal muscle force.13
A recent study by Dunk, et al. (2009), out of the University of Waterloo, evaluated whether the intervertebral joints of the lumbosacral spine approach their end ranges of motion in a seated posture.15 In upright sitting, the L5-S1 intervertebral joint was flexed to more than 60 percent of its total range of motion. In a slouched posture, each of the lower three intervertebral joints approached their total flexion angles. This shows an increased loading of the passive tissues (time-dependent «creep»), which may contribute to low back pain from prolonged sitting.
A study by Reeve, et al. (2009) assessed the thickness of the TrA in various postural positions. Thickness was significantly greater in standing and erect sitting than in a slouched or sway-back standing position.16 The authors concluded that lumbopelvic neutral postures have a positive influence on spinal stability compared to equivalent poor postures.
A study by Claus, et al. (2009) looked at the effect of various postures on regional muscle activity.17 For the deep and superficial fibers of lumbar multifidus muscles, the least muscle activity occurred during a flat posture, which was similar to a slump posture. The most activity occurred in a short lordosis position; there was also more activity in the obliquus internus.
A study by Dolan, et al. (2006) provided evidence that a slouched posture of 5 minutes’ duration can increase reposition error.18 Proprioceptive control is known to be valuable in spinal stability. The fact that reposition error can occur within as little as 5 minutes of «slouched» posture suggests the importance of postural education in decreasing proprioceptive loss and injury.
Om at enkel stretching før leggetid gjør at nattlige kramper blir markant mindre.
http://www.ncbi.nlm.nih.gov/m/pubmed/22341378/
Hele studien her: http://ajp.physiotherapy.asn.au/AJP/vol_58/1/Hallegraeff.pdf
t six weeks, the frequency of nocturnal leg cramps decreased significantly more in the experimental group, mean difference 1.2 cramps per night (95% CI 0.6 to 1.8). The severity of the nocturnal leg cramps had also decreased significantly more in the experimental group than in the control group, mean difference 1.3 cm (95% CI 0.9 to 1.7) on the 10-cm visual analogue scale.
CONCLUSION: Nightly stretching before going to sleep reduces the frequency and severity of nocturnal leg cramps in older adults.
http://origin-ars.els-cdn.com/content/image/1-s2.0-S1836955312700681-gr1.jpg
| What is already known on this topic: Nocturnal leg cramps are common among the elderly, causing pain and sleep disturbance. The medications used to prevent nocturnal leg cramps have variable efficacy and may have substantial side effects. |
| What this study adds: Nightly stretching of the calves and hamstrings reduces the frequency of nocturnal leg cramps in older adults. Nightly stretching also lessens the pain associated with any cramps that continue to occur.
The cause of nocturnal leg cramps is unknown. However, several possible causes and precipitating factors have been hypothesised. Abnormal excitability of motor nerves, perhaps due to electrolyte imbalance, may be a contributing mechanism (Monderer et al 2010). Diuretics, steroids, morphine, and lithium are also reported to cause nocturnal cramps, as can repetitive movements during sport (Butler et al 2002, Kanaan and Sawaya 2001, Monderer et al 2010). Conversely, physical inactivity has been proposed as a cause, with inadequate stretching leading to reduced muscle and tendon length (Monderer et al 2010, Sontag and Wanner 1988). Quinine and hydroquinine are moderately effective in reducing the frequency and severity of nocturnal leg cramps (El-Tawil et al 2010, van Kan et al 2000), perhaps by decreasing the excitability of the motor end plate and thereby increasing the refractory period of a muscle (Vetrugno et al 2007). However, quinine can have important side effects, especially for women, such as: thrombocytopenia, hepatitis, high blood pressure, tinnitus, severe skin rash, and haemolytic uremic syndrome (Aronson 2006, Inan-Arslan et al 2006). Although other medications have been used to treat nocturnal leg cramps such as magnesium, Vitamin B Complex Forte, calcium, and vitamin E, none of these appears to be effective (Anonymous 2007, Daniell 1979). Moreover, stretching techniques can foster a resilient attitude toward recovery in patients with nocturnal leg cramps by promoting a ‘bounce back and move on’ behavioural strategy (Norris et al 2008), because they give patients a strategy to seek immediate relief. Each stretch was performed a total of three times, with 10 seconds of relaxation between each stretch. Stretching of both legs was done within three minutes. Our results showed that six weeks of nightly stretching of the calf and hamstring muscles significantly reduced the frequency and severity of nocturnal leg cramps in older people. The best estimate of the average effect of stretching on the frequency of cramps was a reduction of about one cramp per night. The stretches reduced the severity of the pain that occurred with the nocturnal leg cramps by 1.3 cm on a 10-cm visual analogue scale. |

{kind=link}