Never det meste rundt bindevev: tensegritet, subcutan hud, skinligaments, stretching, ligamenter, nerver, m.m.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2667913/
Fasciae probably hold many of the keys for understanding muscle action and musculoskeletal pain, and maybe of pivotal importance in understanding the basis of acupuncture and a wide range of alternative therapies (Langevin et al. 2001, 2002, 2006a; Langevin & Yandow, 2002; Iatridis et al. 2003). Intriguingly, Langevin et al. (2007) have shown that subtle differences in the way that acupuncture needles are manipulated can change how the cells in fascia respond. The continuum of connective tissue throughout the body, the mechanical role of fascia and the ability of fibroblasts to communicate with each other via gap junctions, mean that fascia is likely to serve as a body-wide mechanosensitive signaling system with an integrating function analogous to that of the nervous system (Langevin et al. 2004; Langevin, 2006). It is indeed a key component of a tensegrity system that operates at various levels throughout the body and which has been considered in detail by Lindsay (2008) in the context of fascia.
Anatomists have long distinguished between superficial and deep fascia (Fig. 1), although to many surgeons, ‘fascia’ is simply ‘deep fascia’. The superficial fascia is traditionally regarded as a layer of areolar connective or adipose tissue immediately beneath the skin, whereas deep fascia is a tougher, dense connective tissue continuous with it.

A diagrammatic representation of a transverse section through the upper part of the leg showing the relative positions of the superficial (SF) and deep fascia (DF) in relation to the skin (S) and muscles. Note how the deep fascia, in association with the bones [tibia (T) and fibula (F)] and intermuscular septa (IS) forms a series of osteofascial compartments housing the extensor, peroneal (PER) and flexor muscles. If pressure builds up within a compartment because of an acute or overuse injury, then the vascular supply to the muscles within it can be compromised and ischaemia results. ANT, anterior compartment; IM, interosseous membrane.
The presence of a significant layer of fat in the superficial fascia is a distinctive human trait (thepanniculus adiposus), compensating for the paucity of body hair. It thus plays an important role in heat insulation. In hairy mammals, the same fascia is typically an areolar tissue that allows the skin to be readily stripped from the underlying tissues (Le Gros Clark, 1945). Where fat is prominent in the superficial fascia (as in man), it may be organized into distinctive layers, or laminae (Johnston & Whillis, 1950), although Gardner et al. (1960) caution that these may sometimes be a characteristic of embalmed cadavers and not evident in the living person. Furthermore, Le Gros Clark (1945) also argues that fascial planes can be artefactually created by dissection. Conversely, however, some layers of deep fascia are more easily defined in fresh than in fixed cadavers (Lytle, 1979).
The superficial fascia conveys blood vessels and nerves to and from the skin and often promotes movement between the integument and underlying structures.
Skin mobility protects both the integument and the structures deep to it from physical damage. Mobility is promoted by multiple sheets of collagen fibres coupled with the presence of elastin (Kawamata et al. 2003). The relative independence of the collagen sheets from each other promotes skin sliding and further stretching is afforded by a re-alignment of collagen fibres within the lamellae. The skin is brought back to its original shape and position by elastic recoil when the deforming forces are removed. As Kawamata et al. (2003)point out, one of the consequences of the movement-promoting characteristics of the superficial fascia is that the blood vessels and nerves within it must run a tortuous route so that they can adapt to an altered position of the skin, relative to the deeper structures.
Although deep fascia elsewhere in the limbs is often not so tightly bound to the skin, nevertheless cutaneous ligaments extending from deep fascia to anchor the integument are much more widespread than generally recognized and serve to resist a wide variety of forces, including gravitational influences (Nash et al. 2004).
According to Bouffard et al. (2008), brief stretching decreases TGF-β1-mediated fibrillogenesis, which may be pertinent to the deployment of manual therapy techniques for reducing the risk of scarring/fibrosis after an injury. As Langevin et al. (2005) point out, such striking cell responses to mechanical load suggest changes in cell signaling, gene expression and cell-matrix adhesion.
In contrast, Schleip et al. (2007) have reported myofibroblasts in the rat lumbar fascia (a dense connective tissue). The cells can contract in vitro andSchleip et al. (2007) speculate that similar contractions in vivo may be strong enough to influence lower back mechanics. Although this is an intriguing suggestion that is worthy of further exploration, it should be noted that tendon cells immunolabel just as strongly for actin stress fibres as do fascial cells and this may be associated with tendon recovery from passive stretch (Ralphs et al. 2002). Finally, the reader should also note that true muscle fibres (both smooth and skeletal) can sometimes be found in fascia. Smooth muscle fibres form the dartos muscle in the superficial fascia of the scrotum and skeletal muscle fibres form the muscles of fascial expression in the superficial fascia of the head and neck.
Consequently, entheses are designed to reduce this stress concentration, and the anatomical adaptations for so doing are evident at the gross, histological and molecular levels. Thus many tendons and ligaments flare out at their attachment site to gain a wide grip on the bone and commonly have fascial expansions linking them with neighbouring structures. Perhaps the best known of these is the bicipital aponeurosis that extends from the tendon of the short head of biceps brachii to encircle the forearm flexor muscles and blend with the antebrachial deep fascia (Fig. 6). Eames et al. (2007) have suggested that this aponeurosis may stabilize the tendon of biceps brachii distally. In doing so, it reduces movement near the enthesis and thus stress concentration at that site.

The bicipital aponeurosis (BA) is a classic example of a fascial expansion which arises from a tendon (T) and dissipates some of the load away from its enthesis (E). It originates from that part of the tendon associated with the short head of biceps brachii (SHB) and blends with the deep fascia (DF) covering the muscles of the forearm. The presence of such an expansion at one end of the muscle only, means that the force transmitted through the proximal and distal tendons cannot be equal. LHB, long head of biceps brachii. Photograph courtesy of S. Milz and E. Kaiser.
Several reports suggest that fascia is richly innervated, and abundant free and encapsulated nerve endings (including Ruffini and Pacinian corpuscles) have been described at a number of sites, including the thoracolumbar fascia, the bicipital aponeurosis and various retinacula (Stilwell, 1957; Tanaka & Ito, 1977; Palmieri et al. 1986; Yahia et al. 1992; Sanchis-Alfonso & Rosello-Sastre, 2000; Stecco et al. 2007a).
Changes in innervation can occur pathologically in fascia, and Sanchis-Alfonso & Rosello-Sastre (2000) report the ingrowth of nociceptive fibres, immunoreactive to substance P, into the lateral knee retinaculum of patients with patello-femoral malignment problems.
Stecco et al. (2008) argue that the innervation of deep fascia should be considered in relation to its association with muscle. They point out, as others have as well (see below in ‘Functions of fascia’) that many muscles transfer their pull to fascial expansions as well as to tendons. By such means, parts of a particular fascia may be tensioned selectively so that a specific pattern of proprioceptors is activated.
It is worth noting therefore that Hagert et al. (2007) distinguish between ligaments at the wrist that are mechanically important yet poorly innervated, and ligaments with a key role in sensory perception that are richly innervated. There is a corresponding histological difference, with the sensory ligaments having more conspicuous loose connective tissue in their outer regions (in which the nerves are located). Comparable studies are not available for deep fascia, although Stecco et al. (2007a) report that the bicipital aponeurosis and the tendinous expansion of pectoralis major are both less heavily innervated than the fascia with which they fuse. Where nerves are abundant in ligaments, blood vessels are also prominent (Hagert et al. 2005). One would anticipate similar findings in deep fascia.
Some of the nerve fibres associated with fascia are adrenergic and likely to be involved in controlling local blood flow, but others may have a proprioceptive role. Curiously, however, Bednar et al. (1995)failed to find any nerve fibres in thoracolumbar fascia taken at surgery from patients with low back pain.
The unyielding character of the deep fascia enables it to serve as a means of containing and separating groups of muscles into relatively well-defined spaces called ‘compartments’.
One of the most influential anatomists of the 20th century, Professor Frederic Wood Jones, coined the term ‘ectoskeleton’ to capture the idea that fascia could serve as a significant site of muscle attachment – a ‘soft tissue skeleton’ complementing that created by the bones themselves (Wood Jones, 1944). It is clearly related to the modern-day concept of ‘myofascia’ that is popular with manual therapists and to the idea of myofascial force transmission within skeletal muscle, i.e. the view that force generated by skeletal muscle fibres is transmitted not only directly to the tendon, but also to connective tissue elements inside and outside the skeletal muscle itself (Huijing et al. 1998; Huijing, 1999).
One can even extend this idea to embrace the concept that agonists and antagonists are mechanically coupled via fascia (Huijing, 2007). Thus Huijing (2007) argues that forces generated within a prime mover may be exerted at the tendon of an antagonistic muscle and indeed that myofascial force transmission can occur between all muscles of a particular limb segment.
Wood Jones (1944) was particularly intrigued by the ectoskeletal function of fascia in the lower limb. He related this to man’s upright stance and thus to the importance of certain muscles gaining a generalized attachment to the lower limb when it is viewed as a whole weight-supporting column, rather than a series of levers promoting movement. He singled out gluteus maximus and tensor fascia latae as examples of muscles that attach predominantly to deep fascia rather than bone (Wood Jones, 1944).
They have argued that a common attachment to the thoracolumbar fascia means that the latter has an important role in integrating load transfer between different regions. In particular, Vleeming et al. (1995) have proposed that gluteus maximus and latissimus dorsi (two of the largest muscles of the body) contribute to co-ordinating the contralateral pendulum like motions of the upper and lower limbs that characterize running or swimming. They suggest that the muscles do so because of a shared attachment to the posterior layer of the thoracolumbar fascia. Others, too, have been attracted by the concept of muscle-integrating properties of fascia. Thus Barker et al. (2007) have argued for a mechanical link between transversus abdominis and movement in the segmental neutral zone of the back, via the thoracolumbar fascia. They feel that the existence of such fascial links gives an anatomical/biomechanical foundation to the practice in manual therapy of recommending exercises that provoke a submaximal contraction of transversus abdominis in the treatment of certain forms of low back pain.
An important function of deep fascia in the limbs is to act as a restraining envelope for muscles lying deep to them. When these muscles contract against a tough, thick and resistant fascia, the thin-walled veins and lymphatics within the muscles are squeezed and their unidirectional valves ensure that blood and lymph are directed towards the heart. Wood Jones (1944) contests that the importance of muscle pumping for venous and lymphatic return is one of the reasons why the deep fascia in the lower limb is generally more prominent than in the upper – because of the distance of the leg and foot below the heart.
In certain regions of the body, fascia has a protective function. Thus, the bicipital aponeurosis (lacertus fibrosus), a fascial expansion arising from the tendon of the short head of biceps brachii (Athwal et al. 2007), protects the underlying vessels. It also has mechanical influences on force transmission and stabilizes the tendon itself distally (Eames et al. 2007).
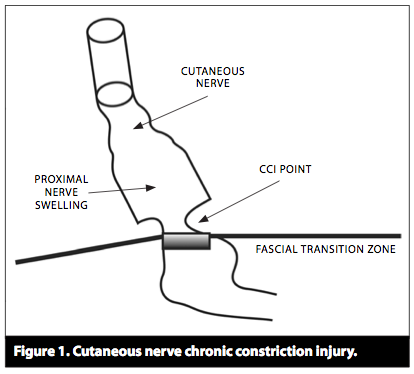
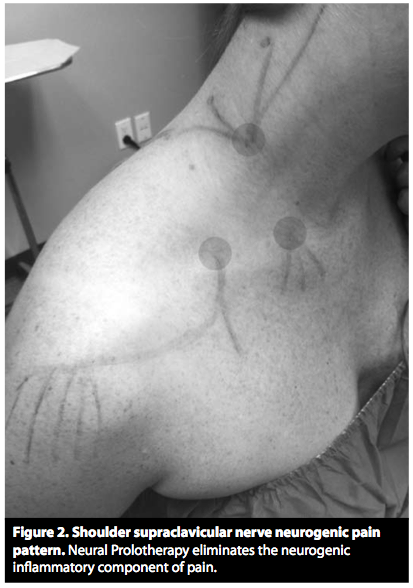
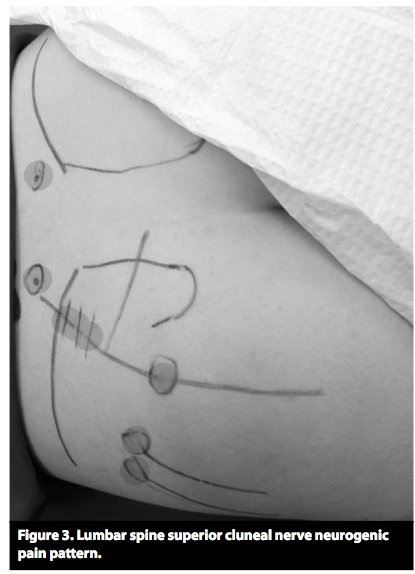
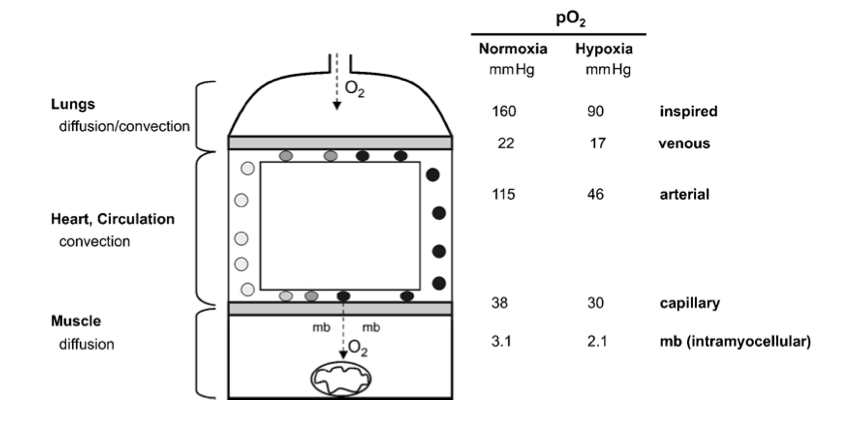





 (
( ) via a hypoxia-mediated increase in their sensitivity to [H+
) via a hypoxia-mediated increase in their sensitivity to [H+ ≥ 150 mmHg) effectively silences this response (
≥ 150 mmHg) effectively silences this response (

 = 150 mmHg) representing central chemoreflex response, and hypoxic (
= 150 mmHg) representing central chemoreflex response, and hypoxic ( = 50 mmHg) representing the addition of central and peripheral chemoreflexes responses. The slope of each isoxic response represents sensitivity of the chemoreflex to CO2. The inflection point at which ventilation starts to increase in response to increasing
= 50 mmHg) representing the addition of central and peripheral chemoreflexes responses. The slope of each isoxic response represents sensitivity of the chemoreflex to CO2. The inflection point at which ventilation starts to increase in response to increasing  is the ventilatory recruitment threshold (VRT), where the chemoreflex neural drive to breathe exceeds a drive threshold and starts to produce an increase in pulmonary ventilation. Ventilation below VRT represents non-chemoreflex drives to breathe and is known as the basal ventilation. The differences in ventilation between isoxic rebreathing lines at any given isocapnic
is the ventilatory recruitment threshold (VRT), where the chemoreflex neural drive to breathe exceeds a drive threshold and starts to produce an increase in pulmonary ventilation. Ventilation below VRT represents non-chemoreflex drives to breathe and is known as the basal ventilation. The differences in ventilation between isoxic rebreathing lines at any given isocapnic  can be used to calculate the hypoxic ventilatory response (indicated by vertical arrows). Note that the choice of isocapnic
can be used to calculate the hypoxic ventilatory response (indicated by vertical arrows). Note that the choice of isocapnic  affects the magnitude of the measured HVR even within the same subject (
affects the magnitude of the measured HVR even within the same subject ( values in the illustrated example. Note also that the magnitude of HVR provides little information about the characteristics of the control of breathing model, as HVR magnitude is dependent on the combination of central and peripheral chemoreflex responses.
values in the illustrated example. Note also that the magnitude of HVR provides little information about the characteristics of the control of breathing model, as HVR magnitude is dependent on the combination of central and peripheral chemoreflex responses. was required to exceed the VRT in highlanders compared to lowlanders. Since both central and peripheral chemoreceptors are actually [H+
was required to exceed the VRT in highlanders compared to lowlanders. Since both central and peripheral chemoreceptors are actually [H+ can be described as follows:
can be described as follows: where [HCO
where [HCO is 40 mmHg and [HCO
is 40 mmHg and [HCO that leads to a reduction in [H
that leads to a reduction in [H /[HCO
/[HCO of 30 mmHg rather than 40 mmHg, as at sea-level. Considering that the highlanders in our study have an adapted acid–base status (
of 30 mmHg rather than 40 mmHg, as at sea-level. Considering that the highlanders in our study have an adapted acid–base status ( relationship in highlanders compared to lowlanders, with the assumption that the chemoreceptor thresholds [H
relationship in highlanders compared to lowlanders, with the assumption that the chemoreceptor thresholds [H


