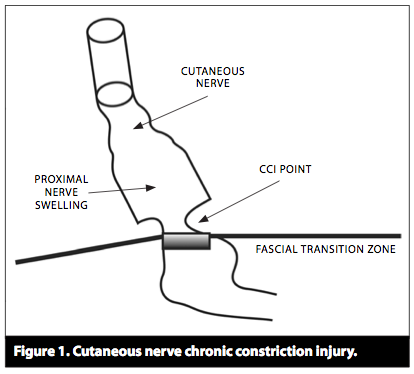
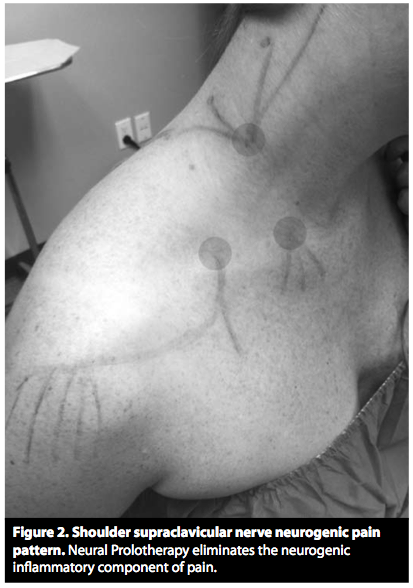
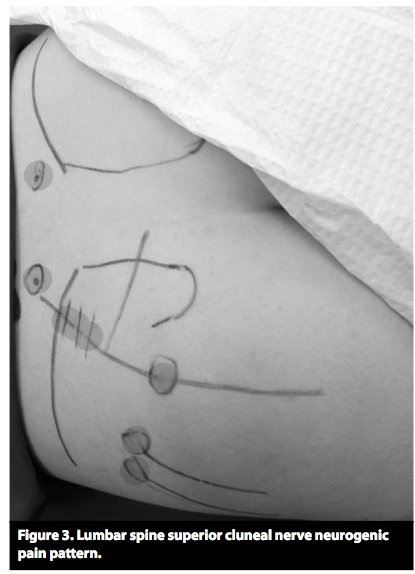
Studie fra 1985 av S.Mense som nevner at det er trykk-sensitive sanseceller i musklene, men at de mest sannsynlig sitter i vevet som omgir muskelen og ikke inni muskelen, siden de ikke reagerer på muskelstrekk eller sammentrekning.
http://m.jp.physoc.org/content/363/1/403.short
Low-threshold pressure-sensitive (l.t.p.) units. Most of these units (ten out of eighteen) responded to touching the receptive field with a soft painter’s brush or a blunt glass rod; they increased their discharge rate upon stimulation with moderate and noxious pressure (Fig. 2). Eight of the l.t.p. units required moderate pressure to be activated;in these cases it was assumed that the mechanosensitive endings were situated more deeply inside the muscle. An observation supporting this assumption was that they did not respond to compression of superficial layers of the muscle with forceps. L.t.p. receptors were relatively insensitive to stretch and contraction; they responded best to forces acting in a direction perpendicular to the long axis of the muscle. Probably, this was due to the fact that many of these receptors were situated in the connective tissue surrounding the tendon and muscle which is not strongly affected by forces building up inside the muscle. (Because of the poor sensitivity of these receptors to the mechanical forces of muscle stretch and contractions,they were not labelled low-threshold mechanosensitive but l.t.p.)
The most frequent type found among group III receptorswas the l.t.p. unit(44%), followed by nociceptive (33%) and contraction-sensitive endings(23%), respectively(Fig.6).
Another feature of l.t.p. units was that they often possessed two receptive fields. In most cases a receptive field with a low mechanical threshold in the distal parts of the muscle or in the tendon was associated with a second receptive field in the proximal or middle third of the muscle, the latter requiring moderate pressure for activation. Out of eighteen l.t.p. units, six(33%) had double receptive fields.
Most of the nociceptive endings had a response behaviour similar to that of cutaneous polymodal nociceptors, in that they responded to both noxious pressure and chemical algesic stimulation.
The physiological significance of the l.t.p. units is not clear. Judging from their response behaviour they would be suited to signal innocuous deformation of muscle and tendon.
In the older literature, there are some reports suggesting the existence of a deep pressure sense in man (Head,Rivers& Sherren, 1905; Lewis & Pochin, 1938; Kellgren & McGowan, 1948), but whether the deep pressure sensations are due to activation of slowly conducting afferent fibres is unknown at present.
 Muskelspenninger helt øverst i nakken bidrar til mange problemer. F.eks. hodepine, spenningsmigrene, kjevespenning, nakkeplager, bevegelsessmerter i nakken, dårlig søvn, m.m.. Spesielt smerter i panna og tinningene har ofte utgangspunkt i området øverst i nakken.
Muskelspenninger helt øverst i nakken bidrar til mange problemer. F.eks. hodepine, spenningsmigrene, kjevespenning, nakkeplager, bevegelsessmerter i nakken, dårlig søvn, m.m.. Spesielt smerter i panna og tinningene har ofte utgangspunkt i området øverst i nakken.