Om hudnære nerver sin rolle i korsryggsmerter. Beskriver spesielt nervene ved L1-2 som går ned til huden i korsryggen og hoftene. Nevner flere interessante diagnosekriterier. Men som de legene og kirurgene forskerne er, har de kun nerveblokkade og kirurgi i som behandlingmuligheter, fullstendig ignorante til mulighetene i manuell behandling. Selv nerveblokkade har 85% av pasientene får mer enn 50% bedring i symptomene, ikke så annerledes enn hva vi forventer med manuell behandling og en intelligent tilnærming til nervesystemet.
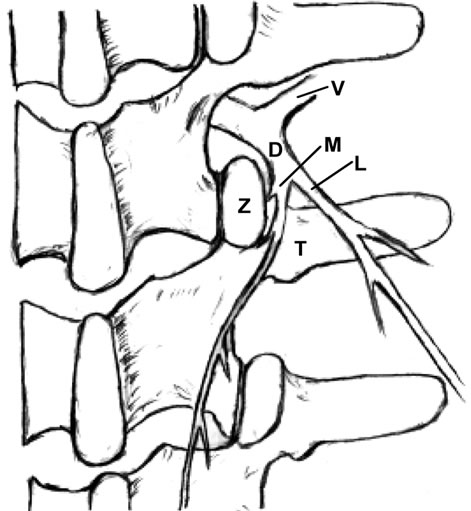
Each spinal dorsal ramus arises from the spinal nerve and then divides into a medial and lateral branch. The medial branch supplies the tissues from the midline to the zygapophysial joint line and innervates two to three adjacent zygapophysial joints and their related soft tissues. The lateral branch innervates the tissues lateral to the zygapophysial joint line.
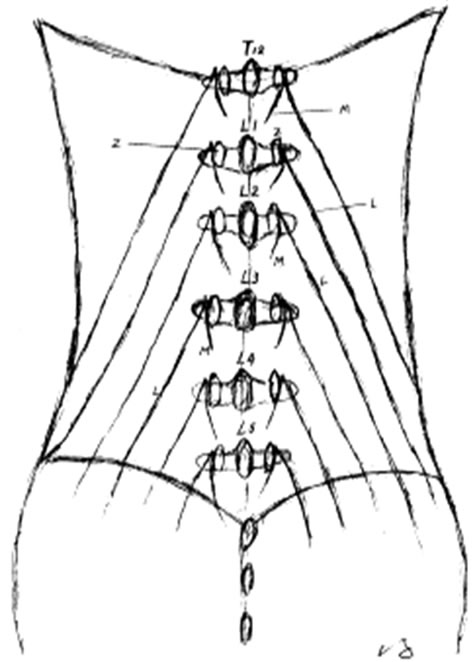
Clinically, L1 and L2 are the most common sites of dorsal rami involvement.
The etiologies of low back pain are numerous. Anatomically, lumbar muscle strain [1,2], lumbar zygapophysial joint syndrome [3-11], instability of the lumbar spine [12], discogenic back pain [1,13], and sacroiliac joint syndrome [2] can cause low back pain. Mechanical pressure on the nerve roots, which may interfere with venous return of the nerve root [14], epidural fibrosis [15], perineural and intraneural fibrosis [16], are additional factors to consider. Additionally, some authors have suggested that the iliolumbar ligament inserting on the lumbar spine is a source of back pain [17].
Anatomically, Bogduk’s work exposed the medial branches of the lumbar spinal dorsal rami as a potential player in low back pain [23-25]. Sihvonen et al. blocked the medial dorsal ramus branch, which resulted in relief of muscle spasms and they suggested that this treatment would aid in improving lumbopelvic rhythm and reducing low back pain [26-29]. These studies supported spinal dorsal ramus as a potential pain generator.
http://file.scirp.org/Html/9-2400120/b3c5afd3-5aa8-4350-9ecc-22fe3e3cc027.jpg
http://file.scirp.org/Html/9-2400120/c6242d6a-bc68-420a-bd92-3c9e7fbafa2d.jpg
The distribution area of each dorsal ramus is characterized by an overlapping multiple segmental innervations, e.g., the L4 zygapophysial joint is innervated by the L3 and L4 medial branches. Therefore, if single dorsal ramus is irritated proximally, a patient could experience pain at the distal site of this nerve distribution (referred pain). This phenomenon mimics radicular pain, for example, a patient with L4-5 herniated disc experiences pain on his dorsal foot. Thus, a local anesthetic injected to the referred pain area will not relieve pain, because the pain is caused by irritation at the proximal dorsal ramus (see below) [36].
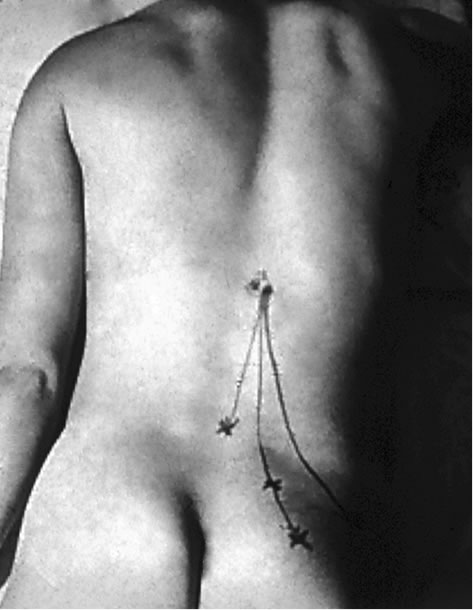
In patients with the spinal dorsal ramus mediated pain, their symptoms usually are on one side and are exacerbated by lumbar extension and/or rotation. This pain may radiate to the ipsilateral low back and buttock region (referred pain) [21,22] (Figure 2). Some patients may present paraspinal muscle spasm (Figure 3(a)). Hyperesthesia may present in the affected dermatome [10,21,22,31,35, 45,46].
http://file.scirp.org/Html/9-2400120/e412d7c3-e859-4500-b82d-396e71f6b638.jpg
The zygapophysial joint line demarcates the distribution of the medial and lateral branches. Pain at between the midline and the zygapophysial joint line or the paraspinal sacroiliac region is caused by on irritated medial branch. Pain lateral to the zygapophysial joint line with radiation to the lateral iliac crest is induced by the lateral branch involvement [19,22]. When the common dorsal ramus is involved, pain will be at the territories of both medial and lateral branches [36,38,47] (Figure 2).
There are some clinical findings in the patient with the spinal dorsal ramus mediated low back pain. The patient usually points to pain at the distal low back (referred pain) [19,22,47]. When the patient bends forward, there is usually a palpable step-off at the affected spinous processes and this is typically two to three segments above the referred pain [47]. There is a palpably widened space and deep tenderness between the spinous processes below the stepoff. With deep palpation of the junction of the same level lateral zygapophysial joint and proximal transverse process, the patient will experience pain and referred pain [19-22,36,47] (Figure 2). Additionally, patients may present an ipsilateral segmental muscle spasm, and a mild scoliosis at the affected vertebral level when the medial branch is involved (Figure 3(a)) [36,47]. If the lateral branch is involved, palpating the longissimus and the iliocostalis muscles can be painful [19-22,35,36,47]. Maigne’s examination techniques are to provoke pain by applying pressure to the lateral aspect of the spinous processes and rubbing the ipislateral facet at the thoracolumbar junction [19-22]. Other findings such as motor, sensory and straight leg raising tests are unremarkable. When the low back pain patient presents pain with radiation below the knee and positive nerve root signs such as loss of sensory or motor function or reflexes in the distribution of the ventral ramus, the ventral ramus involvement (lumbar radiculopathy) should be considered [1].
Any abnormality of the zygapophysial joint such as vertebral malrotation or muscle spasm as well as structural changes of the zygapophysial joint such as subluxation, degeneration, bony proliferation, capsular/ligamentous hypertrophy or fracture can irritate the common dorsal ramus and medial branch, and induce clinical symptoms [18,44,47,48]. Ossification of the mammilloaccessory ligament may cause an entrapment neuropathy and low back pain [18,24,33].
Chen and colleagues [51] dissected the spinal dorsal rami from T12 to the sacrum and conducted biomechanical studies. Their study demonstrated that the L2 dorsal rami bore the greatest stretching force and tensile stress when the specimens were flexed and rotated to the contralateral side.
Spinal dorsal ramus mediated back pain can occur at any level of the human spine [18,21,22,44,52,53]. For low back pain mediated by dorsal ramus, the primary pain is commonly at the thoracolumbar junction [19-22, 38,44]. Within the thoracic region, the coronal orientation of the zygapophysial joints grants spine free rotation. However, this rotation is limited by a rigid rib cage, except at the T10-12 levels because of floating ribs. The upper lumbar facets also have a relative coronal orientation. Therefore, spine rotation is relatively free at the thoracolumbar junction and the greatest shear force occurs at the more mobile upper lumbar segments. This normal spinal movement can cause zygapophysial joint separation or rotation. If these movements occur rapidly or overcome the body’s physiological limit, they can cause stretching tension and irritation to the dorsal ramus, resulting in low back pain [20,40,44,54]. Shao and his colleagues reported that seventy four percent (74%) of the 1263 patients with spinal dorsal ramus mediated low back pain had the pain originating from L1 and/or L2 dorsal ramus [36].
http://file.scirp.org/Html/9-2400120/3fe33e99-0bf8-4fbf-ac3c-867b9de28c6b.jpg
Additionally, Zhou has reported in his retrospective study, that in 41 patients with spinal dorsal ramus mediated lower back pain, after the selective spinal dorsal ramus injection, 84% of these patients received greater than 50% and more than two months of pain reduction [74]. These patients also reported improvement in their daily activities and decrease of their pain medications [74].
Normally, this type of pain originates at L1 or L2 dorsal rami, and the pathogeneses can be multiple factors which irritate the dorsal ramus. The back pain induced by dorsal ramus irritation can occur in the cervical [51] and thoracic spine [50] as well. Therefore, spinal dorsal ramus mediated back pain should be appropriately called “spinal dorsal ramus syndrome (SDRS)”.
The clinical presentations of dorsal ramus mediated back pain and zygapophysial syndrome can be overlapping. However, there are some distinctions. The thoracolumbar junction is the most common site of spinal dorsal ramus mediated back pain [21,22,36], while, zygapophysial joint syndrome commonly occurs at the lower lumbar zygapophysial joints such as L5-S1 and L4-5 [5,6,9,43,57, 58].


 Muskelspenninger helt øverst i nakken bidrar til mange problemer. F.eks. hodepine, spenningsmigrene, kjevespenning, nakkeplager, bevegelsessmerter i nakken, dårlig søvn, m.m.. Spesielt smerter i panna og tinningene har ofte utgangspunkt i området øverst i nakken.
Muskelspenninger helt øverst i nakken bidrar til mange problemer. F.eks. hodepine, spenningsmigrene, kjevespenning, nakkeplager, bevegelsessmerter i nakken, dårlig søvn, m.m.. Spesielt smerter i panna og tinningene har ofte utgangspunkt i området øverst i nakken.



{kind=link}
{kind=link}
{kind=link}
{kind=link}