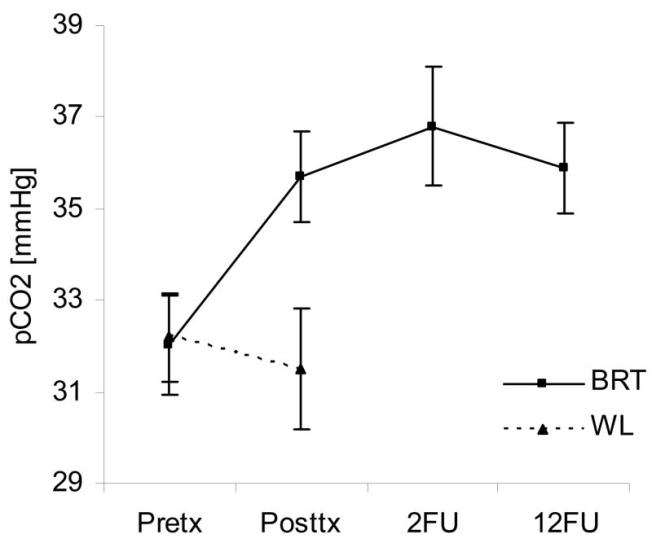
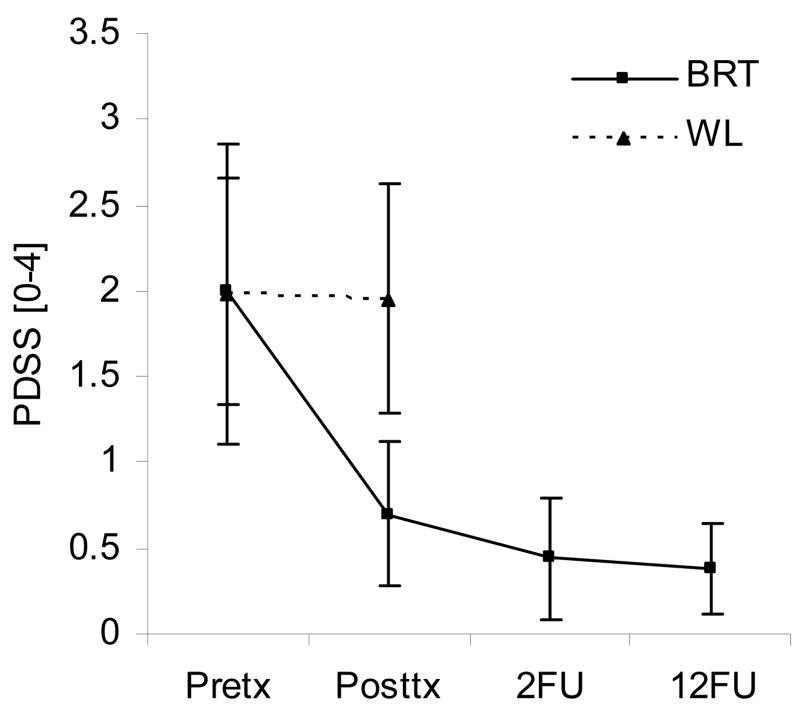
Viktig studie om mekanismen bak hvordan hypoxi gir arrvev (adhesions) i kroppen. Relatert til hyperventilering vil lite CO2 gir hypoxi og sammen med trange blodkar vil de utsatte stedene i kroppen utvikle arrvev mellom muskler og nerver. Nevner hvordan antioksidanter er viktig for å unngå arrvev, spesielt etter operasjoner. Og motsatt, at oksidanter kan skape arrvev fra friskt vev. Nevner også hvordan nitratreaksjoner er med å skaper arrvev, så mulig at CO2 bidrar med å dempe nitratreaksjonene og dermed dempe dannelsen av arrvev. Den viser også at det kan være mulig å få arrvev celler om til å bli normale celler.
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2574925/
«Adhesion fibroblasts exhibit higher TGF-β1 and type I collagen expression as compared to normal peritoneal broblasts. Furthermore, exposure of normal peritoneal fibroblasts to hypoxia results in an irreversible increase in TGF-β1 and type I collagen. We postulated that the mechanism by which hypoxia induced the adhesion phenotype is through the production of superoxide either directly or through the formation of peroxynitrite. »
«Hypoxia treatment resulted in a time-dependent increase in TGF-β1 and type I collagen mRNA levels in both normal peritoneal and adhesion fibroblasts.»
«In contrast, treatment with SOD, to scavenge endogenous superoxide, resulted in a decrease in TGF-β1 and type I collagen expression in adhesion fibroblasts to levels seen in normal peritoneal fibroblasts; no effect on the expression of these molecules was seen in normal peritoneal fibroblasts. »
«In conclusion, hypoxia, through the production of superoxide, causes normal peritoneal fibroblasts to acquire the adhesion phenotype. Scavenging superoxide, even in the presence of hypoxia, prevented the development of the adhesion phenotype. These findings further support the central role of free radicals in the development of adhesions.»
«Postoperative adhesions are a significant source of impaired organ functioning, decreased fertility, bowel obstruction, difficult reoperation, and possibly pain (1,2)
The following popper user interface control may not be accessible. Tab to the next button to revert the control to an accessible version.
«The processes that result in either normal peritoneal tissue repair or the development of adhesions include the migration, proliferation, and/or differentiation of several cell types, among them inflammatory, immune, mesothelial, and fibroblast cells (3)
«Hypoxia, resulting from tissue injury, has been suggested to play an important role in wound healing, and may therefore be a critical factor in the development of postoperative adhesions (4,7)
The following popper user interface control may not be accessible. Tab to the next button to revert the control to an accessible version.
Destroy user interface contrHypoxia is known to trigger the expression of TGF-β1, which consequently increases the expression of extracellular matrix proteins, including type I collagen (4)
The following popper user interface control may not be accessible. Tab to the next button to revert the control to an accessible version.
Destroy user interface contr«Type I collagen synthesis has been shown to be crucially dependent on the availability of molecular oxygen in tissue culture, animal, and human wound healing experiments (8,9)
The following popper user interface control may not be accessible. Tab to the next button to revert the control to an accessible version.
Destroy user interface controlMoreover, exposure of normal peritoneal fibroblasts to hypoxia irreversibly induces TGF-β1 and type I collagen to levels seen in adhesion fibroblasts (4,10)
«Additionally, hypoxia is known to acutely promote superoxide (O2.−) generation from disparate intracellular sources that include xanthine dehydrogenase oxidase (11), mitochondrial electron transport chain (12), and NAD[P]H oxidase (13).
In biological systems, superoxide dismutase (SOD) protects against the deleterious actions of this radical by catalyzing its dismutation to hydrogen peroxide plus oxygen, (14) Whereas SOD breaks down O2.−, xanthine oxidase synthesizes O2.−. Xanthine oxidase appears to be one of the major superoxide-producing enzymes (14)«
«Scavenging superoxide restores both TGF-β1 and type I collagen mRNA levels in adhesion fibroblasts to levels observed in normal peritoneal fibroblasts»
«Normal peritoneal and adhesion fibroblasts treated with super-oxide dismutase, a O2.− scavenging enzyme, exhibited a dose–response decrease (0, 5, 10, 15, and 20 units/ml) in TGF-β1 and type I collagen mRNA levels in adhesion fibroblasts while not effecting normal peritoneal fibroblasts (Figs. 3A and B).»

«Scavenging superoxide during hypoxia exposure protects against the development of the adhesion phenotype»
«Peroxynitrite treatment increased the adhesion phenotype markers, TGF-β1 and type I collagen»
«Adhesion fibroblasts are myofibroblasts, defined as transiently activated fibroblasts exhibiting features intermediate between those of smooth muscle cells and fibroblasts, including the expression of α-SM actin (29,21) and a depleted antioxidant system (22). In normal wound healing, as the wound resolves, the cellularity decreases and the myofibroblasts disappear by apoptosis (23). However, in several pathological cases, including fibrosis, myofibroblastic differentiation persists and causes excessive scarring (24,25).»
«This is further supported by the fact that when O2.− was scavenged, there was in a significant decrease in TGF-β1 and type I collagen in adhesion fibroblasts to levels seen in normal peritoneal fibroblasts. »
«Reactive oxygen species (ROS) are involved in TGF-β-stimulated collagen production in murine embryo fibroblasts (NIH3T3), and the effect of glutathione depletion on TGF-β-stimulated collagen production may be mediated by facilitating ROS signaling (37).»
«Reactive oxygen and nitrogen intermediates control the synthesis of cytokines and growth factors in several in vitro models (40). For instance, they modulate the expression and/or release of monocyte chemoattractant protein-1 (41,42), tumor necrosis factor-α, interleukin (IL)-1 (43,44), IL-8 (45,46), platelet-derived growth factor (47,48), and TGF-β1 (49). «
«Adhesion fibroblasts exhibited a significantly lower level of nitric oxide (NO) and higher protein nitration as compared to normal peritoneal fibroblasts, although there was no difference in the iNOS expression level between the two cell lines (17,50,51). This strongly indicates that adhesion fibroblasts use NO to form ONOO−, and consequently their basal ONOO− levels are higher than normal peritoneal fibroblasts. «
«Thus, treatment with SOD might affect the homeostasis of myofibroblasts by inducing cell death or the phenotypic reversion of myofibroblasts into normal fibroblasts. »
«Our results clearly indicate that hypoxia generated O2.− is a key player in the formation of the adhesion phenotype. This became evident when normal peritoneal fibroblasts were treated with SOD under hypoxic conditions and no change in adhesion markers was seen.»

«In this model, hypoxia-generated O2.− exerts its effect directly by enhancing the expression of TGF-β1, which consequently leads to elevated levels of type I collagen, a hallmark of the adhesion phenotype.»